

Acronyms
Acronym | Description |
|---|---|
AOB | Apparently Occupied Burrows |
AON | Apparently Occupied Nests |
AOS | Apparently Occupied Sites |
AOT | Apparently Occupied Territories |
CEH | Centre for Ecology and Hydrology |
DF | Diversionary Feeding |
DHT | Dunbar Harbour Trust |
DNA | Deoxyribonucleic Acid |
ECON | ECON Ecological Consultancy Ltd |
EEC | European Economic Community |
EIA | Environmental Impact Assessment |
ELC | East Lothian Council |
ELCRS | East Lothian Countryside Ranger Service |
EU | European Union |
FSG | Forth Seabird Group |
FIHG | Forth Islands Heritage Group |
GB | Great Britain |
HES | Historic Environment Scotland |
INNS | Invasive non-native Species |
JNCC | Joint Nature Conservation Committee |
LIFE | The EU’s funding instrument for the environment and climate action |
NVC | National Vegetation Classification |
QGIS | Quantum Geographic Information System (mapping software) |
RIB | Rigid Inflatable Boat |
RSPB | Royal Society for the Protection of Birds |
SMP | Seabird Monitoring Programme |
SPA | Special Protection Areas |
SSER | SSE Renewables |
SSSI | Site of Special Scientific Interest |
SNH | Scottish Natural Heritage |
SPA | Special Protection Area |
SWT | Scottish Wildlife Trust |
UK | United Kingdom |
1. INTRODUCTION
1. INTRODUCTION
- This document has been prepared for Berwick Bank Wind Farm Limited (the Applicant) as part of the derogation case for the Proposed Development. The Report to Inform Appropriate Assessment (RIAA) concluded that compensation was required for Black-legged Kittiwake (hereafter Kittiwake) Rissa tridactyla, Common Guillemot (hereafter Guillemot) Uria aalge, Razorbill Alca torda, and Atlantic Puffin (hereafter Puffin) Fratercula arctica (using the worst case ‘Scoping Approach’). These are collectively referred to as the ‘key species’.
- Compensatory measures for several other species are also identified within this report. These are presented to demonstrate the thoroughness of the compensatory measure selection process undertaken by the Applicant. However, in view of the RIAA’s conclusions the Applicant’s final list of proposed measures only include those targeting the key species.
- Two colony-based measures are proposed as compensatory measures for the Proposed Development and several others have been explored within this report as part of a thorough compensatory measure identification and selection process. It is proposed that the final measures to take forward are:
- Rat eradication and biosecurity to benefit Kittiwake, Guillemot, Razorbill and Puffin nesting at Handa Island;
- Safeguarding the Dunbar Kittiwake colony through wardening and targeted work to reduce human disturbance and other colony-related pressures;
- Background information on how the colony-based compensatory measures were identified and prioritised is detailed in full within the main derogation case for Berwick Bank offshore wind farm.
- This report provides technical detail on each of the colony-based compensatory measures that were taken forward for further consideration and discussion with regulators and Statutory Nature Conservation Bodies (SNCBs) as part of the Applicant’s Seabird Compensation Consultation process.
- The feedback given in response to these consultation meetings was used to prioritise the ‘long list’ of potential compensatory measures, and at the end of this process the measures were divided into three Tiers, which are defined as follows:
- Tier I: Measures that are agreed to be beneficial, were generally viewed positively, and which can be implemented within the short term
- Tier II: Measures that are agreed to be beneficial, but which are difficult to quantify, or which would require a further data-gathering stage prior to implementation.
- Tier III: Measures which were discussed as a part of the consultation process, but which were not progressed due to lack of support from SNCBs and/or regulators, but which may still have the potential to deliver some compensation benefit.
- Tier I measures are implementable. Tier II measures are not currently being progressed but are considered to be beneficial and of potential value as compensation. Tier III measures may be able to provide some compensation benefit but in general have greater difficulties associated with them and have therefore not been progressed (Table 2.1).
- The specific aims of this report are to:
- Demonstrate the range of potential compensatory measures considered and discussed with regulators and SNCBs;
- Provide detail of the evidence base behind each of the potential colony-based compensatory measures;
- Provide information regarding the anticipated compensation benefits, quantifying these wherever possible based on the best available data.
- The evidence base differs for each of the potential compensatory measures. Various types of evidence are presented and discussed that relate to each potential compensatory measure. Information sources include, but are not limited to, scientific studies, site-specific data, historical information, warden’s reports, consultation with experts, and the results of bespoke fieldwork. In each case the potential benefits to both target and non-target species are described. Sources of uncertainty are discussed. Although all the compensatory measures included within this report are without exception additional to current colony management, a section on additionality is included to demonstrate this.
- A brief description of the project and the mechanism for delivery are included to provide a coherent description of how each measure would work. Further detail regarding implementation and monitoring for each of the Tier I measures is provided in full within the Implementation and Monitoring Plan.
- The compensation benefits of the Tier I measures have been quantified using available evidence from a range of sources, which are documented for each measure. Compensation benefits are presented in terms of conservation targets acknowledging that what is predicted may not necessarily be achieved due to the various uncontrollable factors that may potentially impact on wild bird populations (e.g. disease outbreaks, storms, marine heatwaves and other force majeure events). However, in each instance the evidence base is used to generate an estimate of what level of benefit is considered likely on the basis of evidence to be achievable.
- It was not possible to quantify potential compensation benefits for all the Tier II and III measures and therefore this information is only included where sufficient evidence was available for reasonable estimates to be provided.
Table 1.1: Colony-based compensatory measures and their prioritisation ranking.
2. TIER I: RODENT ERADICATION FROM ISLANDS
2. TIER I: RODENT ERADICATION FROM ISLANDS
2.1. BACKGROUND
2.1. BACKGROUND
- Invasive non-native species (INNS) of relevance to seabirds covers a wide range of flora and fauna. Invasive Tree Mallow Malva arborea for example, forms a dense canopy and may displace native vegetation and prevent burrow occupancy by Puffin on some Firth of Forth islands. Moreover, while a range of introduced mammals such as European Rabbit Oryctolagus cuniculus may have a range of indirect ecological effects on seabirds, it is the predatory species that are seen to be the major threat. In the UK, these may include Black Rat Rattus rattus, Brown Rat Rattus norvegicus, Feral Ferret Mustela furo, Feral Cat Felis catus and American Mink Neovison vison. Other native predators such as Stoat Mustela erminea or Red Fox Vulpes vulpes that are not resident but may occasionally colonise islands may also be treated as INNS in this context. Feral Cat, Red Fox and the mustelids predate adult seabirds as well as chicks, with eggs also taken by all but the former species. Rats are seen to be a particular problem for eggs and young chicks, with House Mouse Mus musculus also taking a predatory role in particular circumstances.
- Eradication of INNS from islands has become a focus in the UK following its success in other locations around the world and its role in reducing extinction risk of a number of species including Seychelles Magpie-robin Copsychus sechellarum, Cook’s Petrel Pterodroma cookii and Black-vented Shearwater Puffinus opisthomelas is now widely acknowledged. In the UK, rodent eradication has been particularly successful at improving the breeding success of burrow (and crevice) nesting species such as Manx Shearwater Puffinus puffinus, European Storm-petrel Hydrobates pelagicus and Puffin (Thomas et al. 2017a). Such is the pervasive effect of rodents that their eradication has been termed ‘Island Restoration’ (Thomas et al. 2017a); although, as this may imply restoration of the whole island ecosystem, rather than just the removal of alien invasive species in order to benefit seabirds, this term is not used here.
- The eradication of rodents, from islands was raised and actively recommended by some stakeholders during consultation. Eradication of rodents from islands has the potential to be more successful than removal from mainland seabird colonies where re-colonisation is inevitable, and a rodent-free status cannot feasibly be maintained. By contrast, if eradication from islands is accompanied by effective biosecurity measures, safe nesting habitat for seabirds can result. There are three basic components to a successful campaign; 1) confirmation of presence and the potential scale of the issue, 2) attempted eradication, and 3) the implementation of a biosecurity plan to avoid re-incursions, monitor for re-incursions and to deal with any re-incursions that do occur.
- Flavoured wax blocks that are attractive to rodents are typically deployed to monitor for their presence. This technique shows which areas the rodents may be occupying and will provide some index of abundance. Poison is not used for surveillance purposes due to the risks posed to non-target species (through either primary or secondary poisoning), especially where no rodents are ultimately found. Other surveillance methods include tracking tunnels, cameras and the use of traps, particularly low-maintenance self-setting traps that are deployed for relatively long periods and record the number of target animals killed.
- The eradication phase by contrast typically requires the use of rodenticide, although other methods such as A24 traps, which use a specially designed lure, are under trial. The rodenticide is typically placed around the island by positioning bait stations at intervals in a grid formation. To ensure that all rodents are removed it may be necessary to install rope access routes across cliff faces. The eradication phase is carried out in the winter when rodent numbers are naturally at their lowest, and when natural food supplies are low. This means that there are fewer rodents to catch, and those that do remain are more likely to take the bait in the absence of other food sources. The winter period also avoids any risk of harm to breeding birds, although it does mean that islands have to be accessed during challenging weather conditions.
- Alongside the initial intensive eradication phase, biosecurity measures need to be put in place to ensure the hopefully acquired rodent-free status is maintained. Biosecurity planning involves the identification of risk species and potential ‘pathways’, such as boats, helicopters, visitors, lighthouse boards and construction work. Prevention measures are required to ensure that invasive species are not transported via these potential pathways.
- Surveillance continues thereafter, potentially using the same or similar methods to that deployed at the start of the project. It is essential to have an incursion response plan that comes into force should the reoccurrence of rodents be detected. The quicker the response, the easier it is likely to be to initiate further removal and for this to be successful as only a few animals may be involved (Thomas & Varnham 2016). Incursions can and do happen regularly (T. Chuchyard, Director RSPB Biosecurity Project, pers. comm).
- In the UK, the EU Biosecurity for LIFE Project is underway and aims to eliminate INNS from the 42 islands in the UK that are designated as SPAs by carrying out monitoring to detect INNS and putting biosecurity measures in place to ensure they remain free of invasive species. A number of islands within the Forth Islands SPA form a part of this project, which is funded through a grant that will finish in July 2023. These islands are: Bass Rock, Isle of May, Fidra, Lamb, Craigleith and Inchmickery
- Although the Forth Islands that form part of the Forth Islands SPA are free of rats, it was suggested that the Applicant could investigate and undertake alien invasive species removal and biosecurity at non-SPA islands supporting the key species. This is a compensatory measure that was recommended to the Applicant by the RSPB (A. Dodds RSPB, pers. comm[1].).
2.2. ISLAND SELECTION PROCESS
2.2. ISLAND SELECTION PROCESS
- SSER undertook a review of the non-SPA Forth Islands (see Table A1 in Appendices), which showed two islands where rodent eradication could be undertaken to benefit small breeding colonies of the key species. Landowner and community support is crucial for rodent eradication projects, and of these islands only Inchcolm Island supported colonies of the key species and promising levels of stakeholder support required for rat eradication to be successful. Since Inchcolm is not an SPA, eradication of rats and implementation of biosecurity control measures at this site would not otherwise be undertaken.
- During stakeholder consultation Handa SPA was identified as a potential site for compensation. Although Handa is currently participating in a trial to assess the efficiency of self-resetting A24 traps in a control context around seabird colonies, there is currently no concerted effort underway to eradicate rats from Handa. This work has been undertaken under the EU Biosecurity for LIFE Project, and will end in 2023. The success of the A24 traps is in question, as there are still significant numbers of rats on Handa and at present there is no source of available funding to eradicate them (R. Potter, NE Reserves Manager SWT, pers. comm).
- Handa is designated as an SPA and at the time of designation in 1990 supported populations of European importance for Guillemot (98,686 individuals – 9.3% of the British population and 2.9% of the North Atlantic biogeographic population) and Razorbill (16,394 individuals – 11% of the British population and 1.9% of the Alca torda islandica population). It also supports nationally important colonies of Kittiwake (10,732 pairs, 2.2% of the British population), as well as several hundred Puffins (735 AOB). The most recent counts show there are an estimated 68,524 Guillemots (individuals), 3,749 Kittiwakes (AON), and 5,047 Razorbills (individuals). On the basis that Handa supports national and internationally important colonies of the key species that require compensation from Berwick Bank, it was considered as a suitable island for compensation.
- Concerns were raised following the Applicant’s Compensation Consultation meeting held on 30th March 2022[2] regarding the distance of Handa from the Proposed Development, and the requirement to systematically look at compensation at closer sites before moving onto more distant sites within the SPA network. To address this concern, a systematic search work was carried out to ascertain whether other closer and/or potentially more suitable islands could be identified and potentially investigated.
- The RSPB Islands Database[3] was used to search for all other islands along the east coast of the UK. This dataset was overlain with the Seabird Nesting Counts shapefiles produced by the JNCC[4] to identify islands where the key species were known to be nesting. The aim was to identify any islands supporting breeding colonies of the key species closer to Berwick Bank, which had not previously been identified or considered. Islands from the east coast of the UK were selected as a priority group for examination as it is anticipated that connectivity to the main SPAs would be greater.
- Although there are ~9,000 islands in the UK, only a few (111 in total) are on the east coast, creating a smaller sub-set for further examination. Overlay of JNCC Seabird Nesting Counts data for the key species revealed that only 19 islands on the east coast supported breeding colonies of the key species (see Table A2 in Appendix).
- Once these islands were identified, they were classified according to their suitability for rat eradication. Islands were not considered suitable if they met any of the following criteria:
- They have never supported rats – some islands are naturally rat free as they do not have the habitat to support rats and/or are too far from the mainland for rats to reach.
- SPA islands that have removed/are already removing rats and have biosecurity plans in place.
- Tidally linked islands and/or islands very close to the mainland where risk of incursion is considered prohibitively high.
- Of the 19 islands identified, 15 were designated as SPAs were already free of rats and had biosecurity plans in place. There were four islands remaining: Inchkeith, Inchcolm, the rocky outcrops outside of Dunbar Harbour (used by nesting Kittiwakes), and Marsden Rock.
- Inchcolm has already been identified as a suitable island for rat eradication. Dunbar has also been identified as a non-SPA Kittiwake colony that could benefit from additional resource (see Section 4). Better control of rodents at the main Dunbar Kittiwake colony has been identified as one of the possible improvements that a full-time site warden could implement.
- It is unclear whether there are rodents on Marsden Rock as the cliffs are steep. However, Marsden Rock sits on the beach at low water and would be vulnerable to recolonisation by rats from the mainland, and therefore is not considered a suitable site for rodent eradication.
- Since no other suitable islands near the potentially impacted SPAs were identified where landowners supported rat eradication, the search area was widened to encompass the whole of the UK. This was on the basis that there may be closer sites in the north of Scotland that could offer comparable opportunities for compensation.
- Whilst Handa has the advantage of supporting large colonies of the key species) it is close to the mainland (only 350m) making it vulnerable to further incursion of rats, which can swim distances of up to 2km. Maintaining islands free from rats is easier if they are uninhabited; there are fewer pathways by which rats can recolonise, and therefore less likelihood of incursions. Handa experiences high numbers of visitors during the summer months (9,000 visitors per year), which is a complicating factor in maintaining Handa free from rats in the long term. It was considered possible that an alternative island could offer similar compensation benefits without these complications.
- To explore this the JNCC Nesting Counts data and the RSPB Islands database were used together to identify the top twenty island breeding colonies for each of the key species (see Tables A4-A7 in Appendix). This process was complicated by the structure of the database, which divides the larger islands into several subsites. In the first instance the database was sorted by subsite, and then sense-checked to remove mainland sites. Then counts from all subsites were added together to present counts for whole islands on the basis that this is the most intuitive metric. However, in some instances this was not possible because of the number of subsites and the way in which they had been titled. In most cases the site name took the name of the island, making the process straightforward. However, the larger Scottish Islands (Orkney, Shetland, West Westray, Rousay, Lewis and Skye) were an exception and the main sites were named after cliffs, headlands or other features. Due to the number of sites and subsites on these large islands (hundreds), hand-sorting the counts for each species by island was unrealistic. Furthermore, eradication of rats from these larger islands is considered unfeasible for several reasons: i) sheer cost and scale, ii) the requirement for complete community support (and the inevitable delays this causes in project implementation), and iii) the number of pathways that would need to be controlled to prevent incursion (e.g. regular ferries etc). For these reasons the larger islands named above were removed from the dataset.
- It is understood that the islands supporting the largest colonies will be SPAs, and therefore likely either to be free from rodents or with removal underway and biosecurity plans in place. However, this was considered the most useful method of assessing Handa in the context of other similar islands and to see whether other more suitable islands may be identifiable. It was acknowledged that this process may not identify islands supporting smaller non-SPA colonies which may not have any funding resource to remove rodents, such as Inchcolm. However, the aim was to find a larger site to provide a greater level of compensation. Although it can be argued that the same level of compensation may be achieved by tackling a larger number of smaller sites, and that smaller sites are not encompassed by the Biosecurity for LIFE Project, it is clear from the scale of the Kittiwake, Guillemot and Razorbill colonies present on Handa that several hundred smaller islands supporting colonies of a similar scale to Incholm would be required to provide a similar level of compensation as Handa. For example, Handa supports 16,394 Razorbills, whilst Inchcolm only supports 12 Apparently Occupied Sites (AOS).
- Once the top twenty islands for each of the key species had been identified, the data was examined to positively identify sites that occurred in the top twenty list for more than one of the key species, essentially generating a list of the most important island colonies for the key species when taken together as a group. The results of this process are summarised in Table 3.1.
- A number of islands identified during this process have never supported rats as they are a long way offshore and/or do not support the habitat and resources required to keep rats alive. However, as a part of the Biosecurity for LIFE Project, all SPA islands are being actively monitored to ensure that rats are not transported accidentally (as can and does regularly occur).
- Seven islands were identified as supporting large breeding colonies (i.e. those in the top twenty by number) for all four key species. These islands were: Berneray, Fair Isle, Foula, Isle of May, Mingulay, Noss and Skomer. All of these islands are free of rats, although it is notable that both Foula and Fair Isle have feral cats. However, the impact of the feral cats, particularly on cliff on nesting seabirds, is unknown. Furthermore, the eradication of feral cats from inhabited islands is potentially more contentious than the eradication of rats and, as both islands are inhabited, strong community support would be required. However, it is acknowledged that action at these sites could conceivably benefit a greater number of birds of the key species, but equally there is uncertainty as to what level of impact the cats are having. Rats are more likely to access cliff faces than feral cats. Neither Foula nor Fair Isle is much closer to Berwick Bank than Handa so there is no obvious advantage in terms of proximity.
- Nine islands were identified as supporting large breeding colonies of three of the key species. Of these islands four did not have rats: Skomer, Colonsay, Farne Isles, Flannan Isles and North Rona. Three islands have had or are in the process of having rats removed. These are Rathlin, Great Saltee and the Shiants. Two islands are known to currently have rats and it is understood that the seabirds nesting there would benefit significantly from their removal. These islands were Handa and Lambay (Ireland). Of these, Handa is both closer and supports larger colonies of the key species and on this basis was considered more likely to deliver a greater compensation benefit. The remaining sites in Table 3.1 support large colonies of only two species.
- Although Handa is not one of the major 20 islands which supports Puffin, it does support a small colony of 208 individuals (according to the latest count from SWT carried out in 2021[5]). Inchcolm also has a small Puffin colony (10 birds counted in 2021[6]). It is anticipated that eradication of rats from both islands will enable Puffin to recolonise new habitat. Puffin has undergone significant decline on Handa where it is currently restricted to nesting on Great Stack, which is believed to still be free of rats. On Inchcolm Puffin only nests on the steepest cliffs on the north-west cliffs of the island. Historically Puffins have nested in other areas on both Inchcolm and Handa and could do again if rats were removed. It should be acknowledged that the impacts of rats on burrow-nesting species such as Puffin are likely to be greater than on cliff-nesting species.
- Handa has a known rodent problem and monitoring carried out by SWT over the 2021 season shows that rat numbers are still high even after trials with A24 traps were conducted over the winter of 2020 (Rab Potter, Reserves Manager, pers. comm). Landowner cooperation and community support is of critical importance in getting rat removal efforts underway and in the case of Handa, it is of significant benefit that the landowner is supportive of rat eradication. The owner of Handa also owns some of the adjacent land on the mainland, which could potentially be maintained as a rodent free buffer to reduce the risk of further incursion of rats from the mainland.
- Further consultation would be needed to ascertain whether there are other SPA sites in a similar position as Handa. However, there does not appear to be any island better suited in terms of compensation for the Proposed Development, either in terms of locality, the match between the species assemblage supported and the key species at the Proposed Development, or in terms of the scale of the rat problem.
Table 2.1: Islands supporting large numbers of more than one of the key species (ranked by the number of key species supported). Abbreviations: RA=Razorbill, KI=Kittiwake, GU=Guillemot, PU=Puffin.
Source: Bird counts from JNCC Seabird Nesting Counts (British Isles) - data.gov.uk Data on invasive species from: Invasive mammals (cefas.co.uk)
- Although Handa is not local to the Firth of Forth, it is acceptable to compensate at another site within the same biogeographical region. Seabirds are generally philopatric, however, it is known that some individuals disperse and may not return to their natal colony to breed. For Kittiwake this is thought to be around 15%, with dispersal on occasion covering considerable distances, sometimes involving movements across the North Sea as far away as France, Norway and Sweden (Wernham et al. 2002). On this basis a level of interchange between the east and west coasts of Scotland can be assumed, and therefore removing rats from Handa would constitute compensation in a different, but connected, topographic unit.
- With regards to the other species, Puffin is the least site-faithful and it is believed that 50% of birds will breed away from their natal colony (Wernham et al. 2002). Over the course of a breeding season second- and third-year Puffins have been observed visiting several different colonies sometimes hundreds of km apart, presumably assessing their potential as future breeding sites. On this basis it seems likely that birds from Handa would end up breeding on the east coast of Scotland.
- Guillemot and Razorbill are believed to be highly philopatric, with a Canadian study showing that 83% of young Razorbills and 93% of adult Razorbills returned to the same colony to breed (Lavers et al. 2007). However, even Razorbills can disperse over long distances: a chick ringed on Handa in 1971 was seen four times at the Gannet Islands some 3,219 km away (Lavers et al 2007). Like Puffin, young non-breeding Guillemots will often visit breeding colonies, presumably with a view to returning there later to breed. For example, on the Isle of May, 51 young Guillemots were observed that originated from other UK and Irish colonies (Wernham et al. 2002). Another study from the Isle of May suggests that 25% of Guillemots reared there may have bred elsewhere (Harris et al. 1996). Therefore, some level of connectivity between Handa and colonies on the east coast of Scotland may be reasonably demonstrated for all the key species.
- On this basis implementing compensation at Handa would fulfil the requirement for the project to provide suitable compensatory measures to secure the overall coherence of the national site network.
2.3. INCHCOLM: RAT ERADICATION, BIOSECURITY & COLONY MANAGEMENT
2.3. INCHCOLM: RAT ERADICATION, BIOSECURITY & COLONY MANAGEMENT
2.3.1. Site description
- Inchcolm measures 9 ha in size, reaches 34m at its highest point, and is located only 1 km from the mainland. The island is comprised of two segments (eastern and western), which are linked by a narrow isthmus, at one time covered by the incoming tide, but which has long been built up to form a permanent causeway. The eastern part of the island is hilly rising to approximately 30m above sea level. The western part is somewhat flatter, but gently rises to a similar height at its western extremity where stretches of cliffs can be found. The island is famous for its ancient Abbey and its World War 1 and 2 military fortifications. It has some small sand beaches, but otherwise it is rocky. The grounds of the Abbey and the tourist reception area are landscaped comprising mowed grass, ornamental shrubs and hard standing. The rest of the island is largely dominated by coastal grassland vegetation with small shrubs and trees
- The island is regularly visited by tourist trips to Inchcolm Abbey during the summer months, with up to four employees of Historic Environment Scotland (HES) living on the island during the summer to staff the abbey and shop. The abbey may also be hired as a wedding venue. Inchcolm has no nature conservation designations and no history of management for seabirds. The island is part of the Moray Estate, although HES have responsibility for running and maintaining the abbey.
- Situated within 500m of Inchcolm island there are the two small barren rocky islets of Carr Craig (to the east) and Haystack (to the west) both of which have been important breeding grounds for several species of tern in the past, and in more recent years have hosted important colonies of Great Cormorant and European Shag.
- Inchcolm is known to support a colony of Black Rats. Consequently, it was a surprise when the Forth Seabird Group discovered a Kittiwake colony there in 1991, as it was thought that the rats would discourage nesting seabirds (ENHS Report 1992). Further information on Black rat and the impacts of Black rat on seabird populations is included within the feasibility study (Cain et al., 2022).
- The seabirds nesting on Inchcolm were counted annually by the Forth Seabird Group and are now counted annually by the Forth Islands Heritage Group (FIHG). Only very small numbers nest there with the colony in 2021 numbering 63 AON for Kittiwake, 12 AOS for Razorbill, and 10 Puffins (single birds). Guillemots have been observed on Inchcolm on several occasions (single birds in 2007 and 2008, and 14 individuals in 2014) and it was speculated that a very small number may breed there. However, they have not been sighted on more recent surveys (2015–2021). The number of birds present on Inchcolm is extremely low both relative to other unmanaged non-SPA islands such as Inchkeith and relative to the other rodent-free Forth Islands that are included within the SPA designation, and it is speculated that this could be due to the presence of Black Rat, which is considered to be even more detrimental than Brown Rat as a result of its greater agility and ability to access remote nesting locations.
- Kittiwake numbers on Inchcolm remained relatively stable during the period between 2004–2008 (Figure 2.1) when numbers declined rapidly elsewhere (see Figure 3.1 in Section 3.2.4 for comparison). However, numbers then dropped further between 2014-2018, when numbers have stabilised or even increased elsewhere. The period of relative stability during the mid-noughties (when other sites declined) may be attributable to dietary differences, with Kittiwakes in the Inner Forth (Inchkeith and Inchcolm) taking a higher proportion of clupeids rather than sandeels (Bull et al. 2004). However, it is not clear why numbers declined between 2014-2018 in recent years when they did not elsewhere. Comparison with Inchkeith, another non-SPA site in the Inner Forth, shows that Kittiwake numbers were generally stable between 1996-2014 (unlike other sites in the Forth Islands) with numbers increasing post-2014. It is considered likely that the presence of Black Rat on Inchcolm could be an influential factor in the recent decline of Kittiwake.
- Although Razorbill numbers have risen slightly over the last decade, the colony is still only very small relative to rodent-free islands in the Forth (see Section 2.3.5). Puffin counts are variable, but in recent years are always below 60 individuals, numbering only 10 individuals in 2021 (Forth Islands Heritage Group 2021). However, 28 Puffins were seen by the Forth Island Heritage Group off the north-west cliffs of Inchcolm on 20th July, which is considered to be a more representative number of the population than the official count carried out on 31st May (Forth Islands Heritage Group 2021).
Figure 2.1: Counts of Kittiwake, Puffin, Razorbill and Guillemot taken at Inchcolm (data source Forth Seabird Group website: http://www.forthseabirdgroup.org.uk/index.htm, Forth Island Heritage Group reports 2020 & 2021)
- Anecdotal evidence suggests that numbers of Puffin were previously much higher on Inchcolm with peak numbers between 1992-1995 reaching 100 pairs. In the 1990s the Puffins used to nest in a boulder field in the south-eastern corner of Inchcolm, but now they are restricted to the steeper cliffs on the north-west of the island, where there are now only a few pairs (R. Morris, FIHG pers. comm). It is thought likely that they are confined to the cliffs because of Black Rat. However, even though the few remaining Puffins are nesting in the least accessible part of Inchcolm, Black Rat can still reach them even here (see Section 2.3.3). Although Guillemot does not currently breed on Inchcolm, it is also anticipated that rat eradication would increase the value of the habitat and improve colonisation potential.
- However, the number of large gulls (primarily Lesser Black-backed Gull and Herring Gull) nesting on Inchcolm will also affect auk populations through predation of chicks. The vegetation and ground cover means that conducting accurate counts is challenging, so in 2021 the Forth Islands Heritage Group conducted a large gull census on Inchcolm, which showed that numbers were much higher than previously anticipated with 1641-1789 Lesser Black-backed Gull AOT, 1694-1847 Herring Gull AOT and 7 Great Black-backed Gull AOT. Large gulls were observed nesting in the grounds of the Abbey for the first time due to lack of visitors and maintenance during the 2020 closure due to COVID-19 pandemic lockdown.
- However, any solutions even involving non-lethal control of gulls were not popular with stakeholders (see Section 7.2), who as a general principle did not want to see one species favoured over another (meeting held 8th February 2022). Although maintaining specific areas as ‘gull free’ is standard management practice at other sites, such as the Isle of May, which also supports large numbers of Herring Gulls, this approach is not discussed further beyond briefly flagging its potential value at a non-designated unmanaged site like Inchcolm to aid recovery of auk populations, which are currently very low and therefore potentially vulnerable.
- It is acknowledged that maintaining biosecurity at Inchcolm would be challenging because of the number of visitors to Inchcolm Abbey, and its proximity to the mainland, which means that it is at high risk of colonisation by Brown Rat. However, rodent eradication and biosecurity have been successfully carried out in more challenging locations such as St Agnes and Gugh on the Scilly Isles, which are inhabited. The risk of recolonisation can be minimised through regular surveillance, although the time and effort required to achieve and maintain rodent free status at Inchcolm should not be underestimated.
- Following identification of Inchcolm as a potential island that could benefit from rodent removal, a full feasibility study was commissioned (Cain et al., 2022) of which a summary is provided within this document.
- The feasibility study included: i) a field study (rodent trapping), ii) a nesting habitat assessment, iii) a seabird colony census, and iv) a desk study looking at the assessment of 7 key feasibility criteria described in the UK Rodent Eradication Best Practice Toolkit (Thomas et al. 2017). These are technical feasibility, sustainability, social acceptability, political and legal acceptability, environmental acceptability, capacity and costs.
- The feasibility report concluded that a well-planned eradication programme managed by experienced operators, adequately funded, and supported by the landowner and stakeholders, would result in the eradication of Black Rats from Inchcolm and would improve the habitat for target seabirds[9] to breed more successfully and for colonies to expand.
- Although four of the seven key feasibility criteria were met, further consultation was recommended in relation to three areas: sustainability (that is the required commitment to maintain a comprehensive biosecurity plan), political and legal acceptability, and social acceptability.
- Consultation with a range of local stakeholders was conducted during July and August 2022. The consultation was generally positive with the majority in favour of rat removal. HES have indicated that further stakeholder consultation would be required before this specific measure could be secured, the intention is not to take this measure forward as compensation at this stage. Instead, this measure is being presented as a secondary measure that could be implemented as an adaptive management measure if required post consent.
2.3.2. Field survey of mammalian predators
- A field study was carried out by NBC Environment (working in collaboration with WMIL) to assess the abundance and species of invasive mammalian predators present on Inchcolm. Five nights of trapping was undertaken between 13th-23rd June 2022. A total of 80 traps, 9 cameras, 10 ink tunnels, 10 wax blocks and 6 mice boxes were deployed across Inchcolm (Figure 2.2). A range of survey techniques were employed as this approach has been shown to improve the detection of rats. The survey adopts the methodologies described in the UK Rodent Eradication Best Practice Toolkit (UK Biosecurity for Life) (Thomas et al. 2017). Full details of this work are included in the feasibility study (Cain et al., 2022).
- A total of 29 Black Rats were caught over the five nights of trapping and appeared to be distributed across the island (Figure 2.2). No other mammalian predators were detected. All rats caught in kill traps were necropsied and tissue samples were dispatched to laboratories for DNA analysis including genetic sequencing, rodenticide resistance testing and stable isotope analysis to ascertain the food types contributing to rat diet. Initial examination of stomach contents indicated the presence of egg-shell in at least one sample. Stable isotope analysis of whiskers taken from a sample of the Inchcolm rats showed the rat’s diet comprised a marine high trophic level signature, which could be indicative of seabird predation. However, without taking samples of the target seabirds, this test was unable to differentiate between a seabird food source and another high trophic source, for example a dead seal.
- Abundance (or rat density) is recognised as low (less than 10%), moderate (between 11-25%), high (between 26-50%) and very high over 50% (Moors 1985, King & Forsyth, 2021). The index of rat abundance for Inchcolm was 8 rats per 100 trap nights. This suggests a low rat abundance across the island, but this result may be complicated by the trapping time (summer, June 2022) and the abundance of natural food reducing trapping efficacy. This possibility is confirmed when the trapping results are compared to the index from the tracking tunnels (27 active tunnels per 100 trap nights). This suggests that black rat numbers are moderate to high on Inchcolm ((Cain et al., 2022).
- The DNA analytical results show there were no rodenticide resistance genotypes in the black rats trapped on Inchcolm. This suggests that rats could be controlled/eradicated using first generation or second-generation anticoagulant rodenticides (FGARs or SGARs). Rodenticides such as coumatetralyl or bromadiolone could be used to control/eradicate these populations rather than utilising the more toxic brodifacoum or flocoumafen required for resistant populations.
2.3.3. Nesting Habitat assessment
Site visit
- During the period 13th-23rd June 2022 a site visit was undertaken by NBC/WMIL to estimate potential nest site availability on Inchcolm for each the key species. Estimates of nesting space were obtained by both carrying out a walkover a walkover survey of accessible areas and a circuiting the island by boat to take counts and photograph birds nesting on cliffs. Care was taken not to disturb sensitive nesting areas. The results of this work are presented in full within the feasibility study (Cain et al., 2022).
- Since the survey was conducted during peak breeding period it was not possible to carry out precise study of the physical attributes of the nesting areas. This would need to be carried out outside of the breeding period and would involve gathering additional information on the dimensions of individual rock crevices for puffin and razorbill and the depth and type of superficial soils for burrow nesting species (i.e. Puffin).
- Photographs of habitat of interest were taken and observations of seabird activity were recorded. Unoccupied areas of cliffs and slopes for nesting were noted and these areas were crudely measured with the aid of a laser rangefinder and reference to Ordnance Survey maps and Google Earth imagery. The high tide mark plus a 2m ‘splash zone’ was subtracted from the measured height of the ‘unoccupied’ island feature to provide an estimate of the total area of habitat available for additional nesting. Habitats were photographed and matched to the known nesting preferences of the key species to assess potential habitat availability.
Habitat preferences and nesting densities of the key species
- Guillemot and Razorbill nest in broadly similar habitat types and share colony space (Harris & Wanless, 1987), although Razorbill show a preference for nesting in cavities and crevices as well as nesting on ledges (Hipfner & Dussureault, 2001). The density of nests in Guillemot breeding areas varies between site. Densities as high as 46 pairs m-2 have been recorded (Harris & Wanless, 1987), although a lower density of 20 pairs m-2 is used in these calculations (Harris & Birkhead, 1985). Based on the species composition of other seabird colonies within the Forth Islands (Table 2.4), coupled with regular sightings of Guillemot around Inchcolm, it seems feasible to suggest that Inchcolm has the potential to support a mixture of Guillemots and Razorbills, especially given the proximity of other nearby colonies, such as Inchkeith, which could be a source of potential colonists.
- When assessing habitat availability for Guillemot and Razorbill on Inchcolm, only horizontal rocky ledges were selected. This is a conservative estimate, as short ledges and small rocky crevices and other rocky features can also be used, and inclined ledges can be used if flat areas for egg placement or nest building are present. A crude estimate of the total length of the marked areas (total ledge length) was made. Based on the topography of the cliffs and the width/depth of the ledges as observed during the preliminary site visits, a conservative estimate of an average of 0.3m ledge depth was assumed. This width also aligns well with the published literature, with Birkhead (1977) recording a 0.29 m mean width for ledges occupied by Guillemot. Where photographs showed large, flat rock areas (here referred to as platforms), depth was estimated as 0.6 m. Potential nesting space available for Guillemot and Razorbill (number of pairs) was calculated by measuring the ledge length (in meters), multiplying by the ledge depth (in meters) and finally multiplying by an average nesting density of 20 pairs m-2, as generated from the scientific literature (Harris & Birkhead 1985).
- Kittiwakes favour steep cliffs with horizontal ledges for nesting, often sharing these with other seabirds, particularly Guillemot and Razorbill. Nests are built up on grassy knolls in crevices and on ledges using a mix of grasses and seaweeds. A crude estimate suggests a Kittiwake nesting density of 3 nests per linear metre of suitable ledge is achievable for a healthy colony (Massaro et al. 2001). For kittiwake, only horizontal ledges, recesses and grassy knolls with an estimated depth equal to or greater than 0.3 m visible on the photographs were selected. Estimation of available habitat was calculated by measuring the total length of the marked areas, multiplying by the ledge depth (0.3m) and then multiplying by an average nesting density of 3 pairs m-2.
- Puffins nest colonially on cliff tops and grassy slopes, digging a burrow in which a single egg is laid. Puffin burrow density has been shown to be negatively correlated with distance from the cliff edge and positively correlated with angle of slope. These correlations are biologically significant in that close to the cliff edge, where the angle of slope was steep, breeding success was significantly higher than on adjacent level habitat (Nettleship 1972). On St Kilda burrow densities averaging approximately 0.5 burrows m-2 have been reported (Harris and Rothery, 1988). To reinforce the suitability of the Forth islands habitat for Puffin, on the Isle of May, located 45 km to the east of Inchcolm, only five pairs of puffins were breeding in 1958, while 20 years later, 10,000 pairs were present (Boag et al. 1986).
- For Puffins, estimates are based on the areas of steep grassy banks at the top of cliffs or rocky outcrops only. This is a conservative estimate as boulder fields and shallow inclines can also be used if vegetation is managed to improve access, and on Inchcolm it is known that the boulder field on the south-west of the island was historically used as a nesting area. Potential nesting space for puffin was calculated by multiplying the area of grassy bank on top of cliffs or rocky outcrop (in m2) by an average nesting density of 0.5 pairs m-2 (based on Harris and Rothery 1988).
- In all instances estimates are based on the best available measurements from photographs and on conservative assumptions of ledge length, width, and available nesting space from photographs taken from a vessel. Smaller niches and short sections of ledge, not clearly distinguishable on whole area images, are likely to be available and could provide additional nest habitat.
Estimation of available habitat
- The results of the habitat assessment are summarised in Figure 2.3, which shows the locations of potential areas for colony expansion and Table 2.3 which shows the size and potential capacity of different nesting areas for each of the key species. Photographs showing each of the areas shown on Figure 2.3 and delineating the available ledge species for each of the key species are included in Appendix 1 of this document as Figures A1 to A7.
- The habitat assessments do not take into account predation pressure from large gulls, and competition with other species for breeding space, or other factors such as human disturbance.
- Since there is overlap between habitat that could be used by Kittiwake and by Guillemots and/or Razorbills, the results in Table 2.3 are presented in three ways: i) the total estimated number of pairs (of each species) that could be supported by the available good quality habitat, ii) the estimated number of pairs of each species assuming that Kittiwake outcompete Guillemot/Razorbills for suitable shared good quality habitat, and iii) the estimated number of pairs of each species assuming that Guillemot and/or Razorbill outcompete Kittiwake for suitable shared good quality habitat.
- The habitat assessment indicates that there is capacity on Inchcolm to support an additional 250 pairs of Puffins, giving a total of 255 pairs[10]. Since there is some degree of overlap in habitat between the other three species, assumptions have to be made to produce a broad estimate of how many pairs of each species could occupy the available habitat. Table 2.3 shows the number of pairs of each species that could be accommodated under two scenarios: i) whereby Kittiwake outcompete Guillemot/Razorbill, and ii) whereby Guillemot/Razorbill outcompete Kittiwake. Both scenarios assume that Puffin will not compete with Guillemot/Razorbill or Kittiwake. The feasibility study (Cain et al., 2022)states that the reality is likely to be somewhere between these two scenarios. Therefore, an interim value has been calculated based equal colonisation of the available habitat by both Guillemot/Razorbill and Kittiwake (i.e. a value half way between scenario 1 and scenario 2).
- Based on these assumptions, there is capacity on Inchcolm to support 176 additional pairs of Kittiwake, giving a total of 239 pairs, and to support an additional 210 pairs of Guillemot/Razorbill, giving a total of 222 pairs. Since species specific compensation targets are needed to present the compensation in a metric that is comparable with the predicted impacts of Berwick Bank, then it was necessary to establish what proportion of these birds were likely to be Guillemots and what proportion were likely to be Razorbills.
- The mean ratio of Guillemots to Razorbills within the other Forth Islands (listed in Table 2.4) was examined. Ratios varied between 1.6:1 (Guillemots:Razorbills) on Inchkeith to 21:1 on Lamb. Since Inchkeith is the nearest island to Inchcolm, the ratio from this site was used. This was viewed to be reasonable on the basis that a lower number of Guillemots would be expected as they are not currently established on Inchcolm. Using the Guillemot to Razorbill ratio of 1.6:1 from Inchkeith, the additional 210 pairs were assumed to constitute 129 pairs of Guillemots and 81 pairs of Razorbills. Adding on the number of Razorbills currently present (12 pairs), then the habitat assessment suggests that Inchcolm has the potential to support 93 pairs of Razorbills and 129 pairs of Guillemots.
- However, these numbers are to a certain extent arbitrary and are intended only as an approximate guide so that the compensation benefits can be evaluated in a similar metric to the potential impacts of Berwick Bank.
- It is acknowledged that the benefits of rat eradication are likely to be influenced by a range of site-specific factors, which include (but are not limited to) the following:
- The number of rats present;
- How accessible nesting areas are to rats;
- Current population trends, e.g. if a species is declining rapidly then achieving any measurable improvement is unlikely although it may be possible to slow the rate of decline;
- Large scale climatic events and other unpredictable forces majeure;
- Other factors that may impact negatively on breeding success, e.g. poor weather, floods, storms; and
- How easy it is to remove rats and keep the site rat free (e.g. are rope access routes around cliffs required? Is it possible to remove all the rats or is there a significant risk that eradication will be unsuccessful? Is the risk of reinvasion high.)
2.3.4. Project description
- The project to eradicate Black Rat from Inchcolm would be developed in accordance with the UK Rodent Eradication Best Practice Toolkit (Thomas et al. 2017a). A brief outline of the work is included here, and further detail is provided within the Implementation and Monitoring Plan. Further information on the approach to developing a Biosecurity Plan, incorporating information on how incursion of invasive mammalian predators will be prevented, surveillance to monitor for incursion, and incursion response plans should an incursion occur are all included within the Implementation and Monitoring Plan.
- Stakeholder engagement with local groups including HES, the Forth Seabird Group, and the Forth Islands Heritage Group, has been positive and details of a proposed Communication and Engagement Strategy going forward are included in the Implementation and Monitoring Plan. The outputs of the feasibility study also indicate that the project is both technically feasible and would improve seabird breeding success. Although there is a high incursion risk, this can be minimised through positive stakeholder engagement, most notably with the local ferry operator and HES, as well as implementation of a Biosecurity Plan. All funding for this work would be supplied by the Applicant, as outlined in the Implementation and Monitoring Plan.
- As is standard practice, rodent eradication would be carried out during the winter months. The details of rodent removal methods and indicative programme are included within the Implementation and Monitoring Plan.
- A monitoring programme to undertake seabird counts and to monitor productivity would be required to study how key species respond following rodent eradication. Details are included within the Implementation and Monitoring Plan.
- Since Inchcolm is not an SPA and is not covered by any nature conservation designations, there is currently no resource to deal with any work on seabirds. There is clear potential for Applicant funded staff involved in the surveillance and monitoring phase of the project to also tackle any other minor issues that would be part of routine colony management at designated sites, and which are likely to improve seabird breeding success.
- During the stakeholder consultation process undertaken to develop the colony measures it was identified that Inchcolm currently supports Tree Mallow in places around the Abbey gardens. It is recommended that this could be removed to prevent the issues experienced on the other Forth Islands where this invasive species has colonised and grown to such an extent that Puffins are obstructed from entering and leaving their burrows. Additional vegetation management in Puffin nesting areas could also be beneficial. The accumulation of plastic litter on the beaches was also raised. Although HES maintain the Abbey grounds there is currently no mechanism to remove plastic from the rest of the island. It was suggested that an annual plastic pick-up could be enough to maintain Inchcolm in a better condition for both seabirds and also Grey Seal, which use the beach as a breeding site and are vulnerable to entanglement in plastic litter and old fishing nets. Any additional measures to improve seabird nesting habitat and the wider habitats on Inchcolm is detailed in in the Implementation and Monitoring Plan.
- Putting measures in place to reduce human disturbance, particularly during the first half of each calendar year, would also be of benefit to nesting birds, and it would be beneficial if guidelines to minimise disturbance were developed with HES as a component of this project
2.3.5. Conservation targets
- Establishing conservation targets for Inchcolm is problematic as the site has always supported Black Rat for as long as seabird counts have been conducted. Therefore, any predictions of how the bird populations would respond to rat eradication are to a certain extent speculative.
- Maximum counts represent the maximum number of birds that are known to have nested on Inchcolm. For Puffin this is thought to be around 200 birds during the period 1992-1995 (R. Morris pers. comm.). For Kittiwake it is 190 AON (1995), and for Razorbill 21 AOS (2021). However, since Black Rat has always been present, and numbers of birds in general are very low relative to other islands in the Firth of Forth (see Table 2.4), they are not considered representative of what the site may support without Black Rat.
Table 2.3: Numbers of the key species nesting on different Forth Islands.
Data sources: FIHG 2021, Forth Seabird Group 2017, 2018, 2019, 2020 & 2021. Note The size of colonies nesting on Inchcolm are notably low.
- The results of rat eradication projects at other islands can provide helpful information with regards to establishing potential benefits. Examination of data from 12 islands from which rodents have been removed shows increases in the numbers of nesting birds at 10 of these islands (Thomas et al. 2017a, Table 2.5). The two sites where increases were not recorded were Looe Island in Cornwall and Inchgarvie in the Firth of Forth. Both of these sites have had rat re-invasions. The rats have been removed a second time from Looe Island but the seabird population trends are uncertain. Anecdotal rat sightings have been reported from Inchgarvie, and at present there appears to be no monitoring and no resource to carry out further rat removals (Thomas et al. 2017a). Following the production of this review, rats have recently recolonised Handa.
Table 2.4: UK & Crown Dependency rodent-eradication attempts. (Reproduced in full from Thomas et al. 2017a
- At five of the 12 islands, previously lost seabird species recolonised after rodent eradication including Puffin at one site, European Storm-petrel at three sites and Manx Shearwater at one site. At three islands there have been very substantial increases in numbers of Puffins and Manx Shearwaters: on Handa Puffins increased rapidly in the five years following rat eradication, at Ramsey Manx Shearwaters have increased more than five-fold, and at Lundy Manx Shearwater has increased 10-fold and Puffin numbers have increased from five birds to more than 200 pairs (Thomas et al. 2017a).
- Other ecosystem benefits include increases in passerines and ducks as well as other species such as Pygmy Shrew Sorex minutus, Slow Worm Anguis fragilis, Common Lizard Zootoca vivipara, Lesser White-toothed Shrew Crocidura suaveolens, Bank Vole Myodes glareolus and even moths (Thomas et al. 2017a). On this basis, the eradication of rodents from islands is clearly of very significant benefit to seabirds, other avian species, and the whole ecosystem. However, there is an element of uncertainty in predicting recolonisation, and which species will benefit and how rapidly.
- The restoration potential of Inchcolm is very clear both when viewed in the context of the other Forth Islands (Table 2.4) and when the benefits of rat eradication achieved by other eradication programmes are taken into account. On this basis, and on the grounds that data gathered in the absence of rats is lacking, the number of birds that might occupy Inchcolm based on the habitat assessment appears to be the most suitable measure available to use as a basis for conservation targets. However, the results of the habitat assessment are discussed below by species in relation to historical counts, the size of colonies present on the other Forth Islands, and the results of rat eradication programmes conducted elsewhere to ensure that the conservation targets are sense checked and appear reasonable and achievable.
Puffin
- Currently there are five pairs of Puffin nesting on Inchcolm, where they are restricted to the steepest cliffs on the north west of the island. During the period 1992-1995 around 200 Puffin are known to have nested on Inchcolm (R. Morris pers. comm.), utilising a boulder field on the south of the island. Many of the Forth Islands support substantial Puffin colonies, with the nearest on Inchkeith numbering 2178 AOB. Even accounting for Inchkeith’s larger size, it is clear that Puffin numbers on Inchcolm are severely depressed.
- The benefits of rodent eradication for Puffin have been observed at Lundy, Ailsa Craig, Canna, and the Shiant Isles. At Lundy, Puffin have increased by 369% (Brooker et al. 2018), rising from 13 birds in 2000 to 848 in 2021[11]. At Ailsa Craig they have recolonised since rat eradication in 1994 and numbered 130 nests in 2015[12]. On the Shiant Isles productivity increased from 0.66 to 0.79 over the two years following rat eradication and numbers have increased significantly[13]. At Handa, Puffin numbers increased from 472 Puffin in 1996 to 735 Puffins in 1997. This equates to a 56% increase and a gain of ~44 Puffin per year (see Section 3.4.5).
- At Canna, Puffin numbers were low before rat eradication, and they were confined to two virtually inaccessible stacks, making counting them impossible. However, a count of rafting Puffin in 1999 gave 1190 individuals, whilst in 2016, 2050 were counted suggesting a percentage increase in the region of a 70% (although the imprecise nature of counting rafting birds is acknowledged). After rat eradication in 2005/06 Puffin recolonised the main island and have spread to several sites along the north coast of Canna itself.
- The only site where Puffin have been present and not increased significantly following rodent eradication has been Puffin Island, Anglesey. However, there are some indications that even here a slow recovery is underway with an increase from 8 pairs in 2010 to 29 birds in 2018 (still a 263% increase).
- Nationally, Puffin numbers have increased in some locations and declined at others. However, on the grounds that many of the increases described above have been substantial (well in excess of national trends) then it is clear that rat eradication is a powerful measure for Puffin.
- The habitat assessment indicates that Inchcolm has the potential to support an estimated 255 pairs of Puffin. This seems very feasible when compared with the large numbers of Puffins that currently nest on other similar sized islands within the Firth of Forth (e.g. Craigleith, Lamb, Fidra, Inchkeith - see Table 2.4). Inchcolm supported 200 Puffin historically even when rats were present. On this basis the conservation target of 255 pairs of Puffin seems reasonable and achievable based on the proven success of other rat eradication projects for this species.
- It is acknowledged that predation from large gulls and Peregrine Falcon may inhibit recovery and it is suggested that artificial ground cover could be used to reduce avian predation. Previous studies show that Guillemots nesting in areas with artificial cover installed over the cliff tops produced twice as many eggs (Parish and Paine 1996). Diversionary feeding of the Peregrine Falcon pair could also provide an acceptable means of temporarily reducing predation pressure to facilitate the recovery of auk (and also Kittiwake) populations if recovery is slow, although this would require specific investigation (see Section 6).
- Vegetation management, comprising reduction in height and density of grasses and shrubs and loosening of soils on tops of steep slopes, could be adopted prior to the start of the nesting season to optimise conditions and improve access for Puffin. Other practical measures to assist population recovery could include controlling human disturbance around areas of suitable habitat (including the boulder field).
Razorbill
- Currently there are 12 pairs of Razorbills nesting on Inchcolm, where they are restricted to the steepest cliffs on the north west of the island. Razorbill numbers have been very gradually increasing on Inchcolm with maximum counts of 21 AOS recorded in 2020. However, in comparison with the other rat-free Forth Islands, numbers at Inchcolm are extremely low (Table 3.2). Looking at number of AOS per hectare for each island, Inchcolm scores lowest at only one Razorbill per hectare. This figure is highest for The Isle of May at 108 AOS per hectare. Fidra has the lowest number of AOS per hectare of the rat-free Forth islands at 13 AOS per hectare. Since current numbers for Inchcolm are so low, and Razorbills often nest in locations that are accessible to rodents, it seems reasonable to assume that they are significantly impacted by the rats.
- There is less available information on the benefits of rodent eradication and biosecurity for Razorbill, although this will change as more data emerges. However, since Razorbills nest either on lower cliff ledges or among boulders at the bottom of cliffs, then significant improvements to breeding success following rodent eradication are anticipated. Indeed, there is evidence of increases in Razorbill numbers at five islands where they have been monitored following rodent eradiation, namely Canna, Lundy, Ramsey, the Shiants and Ailsa Craig (Table 2.5, Brooker et al. 2018[14], RSPB News from the Rock 2021). However, Razorbill has been increasing nationally and so caution is required in interpreting this information.
- At Lundy, Razorbills increased from 950 in 2000 to 3,533 in 2021, an increase of 272%[15]. When rats were eradicated from Canna over the winter of 2005/06 Razorbill numbers initially increased sharply but then levelled off. However, eggs appeared in areas that had been clear of nesting for several years. Since Razorbill numbers had been declining, rat eradication had the effect of halting the decline (Luxmoore et al 2019). More recently (as the prey supply offshore has ameliorated) there have been big increases in Razorbills nesting on Canna, with numbers doubling between 2016 and 2019. They are now at their highest level since 1995 (The Seabird Group 2019). Comparison of all island Razorbill counts on Handa from 1997 (the year of rat eradication) and 2001 show that Razorbills increased from 15,573 to 17,042, an increase of 1,469 individuals (or 9%) (see Sections 2.4.2 and 2.4.5).
- The habitat assessment indicates that Inchcolm has the nesting space to support an additional 81 pairs of Razorbills (a total of 93 pairs), assuming a mixed Guillemot and Razorbill colony. This is only an approximation and there may be greater numbers of Razorbill if Guillemot are either slow to recolonise or do not recolonise. Habitat assessment indicates that capacity for Guillemot/Razorbill combined is 420 pairs, although some of this habitat would also be suitable for Kittiwake. On this basis achieving a conservation target of 93 pairs seems very feasible in the context of the Razorbill colonies on the other Forth Islands, and acknowledging the benefits achieved elsewhere e.g. Lundy and Canna.
- The recommendations described above to aid Puffin recovery would also help Razorbill populations to increase.
Guillemot
- Although there are no Guillemots on Inchcolm, it is a species that is observed (single birds in 2007 and 2008, and 14 individuals in 2014 – data from annual Forth Seabird Group reports). The habitat assessment (Cain et al., 2022) concludes that Guillemot could reasonably be expected to colonise Inchcolm following rat removal, based on habitat available and the proximity of other islands and sites supporting large breeding colonies. Since other rat free seabird islands within the Firth of Forth support both Guillemots and Razorbill (Table 2.4), it is considered likely, based on the habitat, that Inchcolm could similarly support colonies of both species if nesting conditions were more favourable. Habitat assessment indicated that Inchcolm has the capacity to support 129 pairs of Guillemots. This is considered feasible in view of the numbers of Guillemots supported elsewhere, including on the nearby island of Inchkeith, which could act as a source of potential colonists.
- Rat eradication programmes at other sites have proven successful for Guillemot. For example, following eradication of rats from Lundy, Guillemot numbers increased by 51% (Brooker et al. 2018). At Canna rat eradication had the effect of slowing the rate of decline (Luxmoore et al. 2019), although more recently Guillemots have increased reaching their highest number since 2004 (The Seabird Group 2019).
- Colonisation prospects could be further improved by using decoys and playbacks. Other studies have shown these techniques to be highly effective Parker et al. (2007). Breeding Guillemots were lost from a colony in California following an oil spill in 1986 and did not naturally recolonise over the following eight years. During this period Guillemots were not observed on site. In January 1996 decoys, playbacks and mirrors were installed to attract Guillemots. Following installation Guillemots were observed on site, with 6 pairs breeding in June 1996. By 2004 numbers had increased to 190 pairs (Parker et al. 2007).
- In some seabird species, white paint has been used to simulate guano at potential breeding sites (Gummer, 2003; Sawyer and Fogle, 2013). This could be used to encourage colonisation by all of the auk species, potentially alongside the use of decoys and playbacks, with the aim of increasing colonisation rates following rat eradication.
Kittiwake
- Currently there are 63 pairs of Kittiwake nesting on Inchcolm (FIHG 2021), where they are restricted to the steepest cliffs on the north-west of the island. Although Kittiwakes generally select steep cliff faces for nesting to avoid ground predator, the cliffs on Inchcolm are not that steep, so it is likely that nesting areas will be accessible to rats. Maximum Kittiwake counts on Inchcolm date from 1995 when 190 AON were present (despite the presence of Black rat).
- Although rat eradication projects have not focussed on this species previously many Kittiwakes nest at the top of cliffs and need relatively substantial ledges on which to build nests. The nests themselves also add extra material and structure to the cliff face which may make it easier for rats to climb. Data from Canna shows that Kittiwake numbers have risen significantly since rat eradication in 2005, at a time when Kittiwake was experiencing significant decline elsewhere in Scotland (see Figure 3.1 and also Figure 2.4).
- The habitat assessment shows that Inchcolm has the habitat to support 176 additional pairs of Kittiwake, totalling 239 pairs when adding the 63 pairs currently present (FIHG 2021). This number is relatively small in comparison to some of the other Forth islands (Table 2.4) but is considered reasonable and achievable as a conservation target: much of Inchcolm is relatively low lying and it does not support the same quantity of steep cliff faces as many of the other Forth Islands. However, higher historic counts confirm that nonetheless Inchcolm is capable of supporting a larger Kittiwake colony.
Summary
- Conservation targets for Inchcolm are expressed in Table 2.6 both as the total number of birds that would be generated throughout the 35 year operational lifespan of the Proposed Development, assuming that conservation targets are met, and also as a number of birds that would be provided per year. The numbers are based on the assumptions that have been described above in sections 2.3.3 and 2.3.5. In reality, it is anticipated that it would take considerably less than 35 years to achieve conservation targets – seabird populations have expanded rapidly over 5-10 years on many islands where rats have been eradicated. However, the metric is useful in enabling comparison with predicted annual mortality from the wind farm.
- Although the habitat assessment provides a measure of how many nesting birds Inchcolm could accommodate if conditions were more favourable, it is acknowledged that many factors (including predation pressure from large gulls, and competition with other species for breeding space, or other factors such as human disturbance )may impact on recovery, and as such there is uncertainty associated with whether the conservation targets can be achieved. Conversely, recovery may be enhanced by the measures described in the previous section, such as the use of decoys and playback for Guillemot and artificial ground cover to reduce avian predation.
- Whilst the conservation targets indicate what could potentially be achieved at Inchcolm, they do not measure how improved growth and colony productivity could positively contribute towards seabird populations within the Forth Islands. If the conservation targets are achieved within the next 10 years then it is conceivable that the expanded colonies would produce reasonable numbers of fledged chicks per year, and that this would impact positively on the Forth Islands colonies.
- For example, assuming a productivity value of 0.69 chicks per pair (JNCC 2021) an additional 250 pairs of Puffins could produce an additional 146 fledged chicks per year. Using the mean annual survival rates presented in Horwill & Robinson (2015) and assuming a first breeding age of seven years (Harris & Wanless 2011) this would result in the addition of 72 adults per year being added to the Forth Islands population.
- Similarly, assuming a productivity value of 0.57 chicks per pair (JNCC 2021) an additional 81 pairs of Razorbill could produce an additional 46 fledged chicks per year. Using the mean annual survival rates presented in Horwill & Robinson (2015) and assuming a first breeding age of 4 years (Lavers et al. 2008) this would ultimately result in the addition of 21 adults per year to the Forth Islands population.
- Assuming that colonisation of Inchcolm by Guillemot is achieved and based on a potential 129 pairs of Guillemot occupying the available habitat on Inchcolm and a productivity value of 0.70 chicks per pair (JNCC 2021), then 90 fledged chicks would be produced per year. Using the mean annual survival rates presented in Horwill & Robinson (2015) and assuming a first breeding age of 6.6 years (Harris et al. 2016) this would ultimately result in the addition of 30 adults per year to the Forth Islands population.
- Assuming an additional 176 pairs of Kittiwakes nest on Inchcolm, and assuming a productivity of 0.83 chicks per pair (JNCC 2021), then an additional 146 fledged chicks would be produced per year. Using the mean annual survival rates presented in Horwill & Robinson (2015) and assuming a first breeding age of 4 years (Coulson 2011) this would ultimately result in the addition of 72 adults per year to the Forth Islands population.
- Although these potential benefits are acknowledged, they have not been incorporated within the main compensation benefits tables as they indicate what may be achievable over the longer term following a recovery phase. Currently bird numbers on Inchcolm are very low, and it is acknowledged that recovery may take time. Although it is possible that rats may be the principal reason why bird numbers are so low, and their removal may facilitate a rapid recovery, the uncertainty in this situation is acknowledged and a precautionary approach adopted in the calculation of the potential compensation benefits.
Table 2.5: Preliminary conservation targets and associated increases for each key species on Inchcolm island. All numbers are expressed as single birds.
- Whilst habitat availability provides a theoretical estimate of the numbers of birds that could physically occupy Inchcolm, it does not capture the influence of the factors listed above. Rat activity on Inchcolm is classified as ‘moderate to high’, the nesting areas are accessible to rats, and on this basis significant benefits of rat eradication are anticipated.
- Although Inchcolm supports large colonies of Herring Gull and Lesser Black-backed Gull, which may suppress numbers of Kittiwakes and auks, it is notable that the Isle of May also supports extensive large gull colonies, as does Fidra and Craigleith. However, Inchcolm is located some distance down the Forth Estuary, so it is possible that it may always be less desirable habitat for species that consistently forage offshore, such as Puffin.
2.3.6. Benefits to other species
- Removing rats from Inchcolm could also benefit other birds most notably Fulmar (currently 174 AOS), Shag (27 AON) and Eider (122 AON) (Forth Islands Heritage Group 2021). Other ground-nesting birds known to occur on Inchcolm that would significantly benefit from rat eradication include Mallard Anas platyrhynchos, Shelduck Tadorna tadorna, Oystercatcher Haematopus ostralegus and Wood Pigeon Columba palumbus (Morris 2003). Other species that may benefit to a lesser extent include Rock Dove Columbia livia, and Jackdaw Corvus monedula. A study of islands with and without rats showed that islands with invasive rats had reduced species richness of passerines (Tabak et al. 2015). Therefore, rat eradiation may also offer benefits to the passerine species that are routinely observed on Inchcolm, which include Pied Wagtail Motacilla alba, Dunnock Prunella modularis, European Robin Erithacus rubecula, Blackbird Turdus merula, Chaffinch Fringilla coelebs and Starling Sturnus vulgaris (Morris 2003).
- Previously terns are known to have nested in the local area, either on Inchcolm itself or more commonly on the nearby islet of Carr Craig. Common and Arctic terns are also recorded as nesting on Swallow Craig, a small rocky islet in Inchcolm’s harbour. Previously Common Tern Sterna hirundo, Roseate Tern Sterna dougallii and Sandwich Tern Thalasseus sandvicensis have nested on Inchcolm. It is possible that Common or Arctic Terns could return to Inchcolm, potentially recolonising from the Isle of May, if conditions were more favourable. However, the chances of this would be much improved if there was a possibility of maintaining some areas as gull free.
- Inchcolm is known to support a number of plants typical of coastal grassland and sand dune habitats. These include Sea Rocket Cakile maritima and various Atriplex species (Morris 2003). Both of these plant species increased significantly following the 1997 rat eradication from Handa (see Section 2.4.2), and it is possible that rodent eradication from Inchcolm could benefit both these, and other plant species.
2.3.7. Timescale and mechanism for delivery
- Full details of how the project could be delivered are included within the Implementation and Monitoring Plan, and only a brief summary is included here.
- It is anticipated that the initial rat eradication phase would be undertaken by an eradication specialist during the winter months, with surveillance and seabird monitoring conducted by a member of staff funded by the Applicant, who would also be responsible for implementing incursion response plans should an incursion occur.
- Further stakeholder consultation would be required before this specific measure could be implemented, the intention is not to take this measure forward as compensation at this stage. Instead, this measure is being presented as a secondary measure that could be implemented as an adaptive management measure if required post consent.
2.3.8. Additionality & uncertainty
- Inchcolm does not have any type of nature conservation designations. Currently annual seabird counts are conducted by FIHG, a local volunteer group. There are no plans from any conservation bodies to carry out rat eradication from non-SPA islands. Therefore, all the work proposed is additional; there is no other known mechanism whereby any sort of seabird work at Inchcolm would otherwise be undertaken.
- The factors that may affect the success of rat eradication are listed in above. It is also acknowledged that the positive response of seabird populations following rodent eradication varies significantly between sites and is difficult to predict, especially with regards to recolonisation of species.
- In some locations, birds have not responded as predicted following rat eradication. For example, it was thought that Manx Shearwaters would increase on Canna and Sanday following rodent eradication. These islands are only 4 km from Rum, which holds a third of the global population of Manx Shearwater. However, Manx Shearwater numbers have remained low, despite the increases amongst other species. Similarly, the timescale in which existing species increase is also very variable between sites.
- Although Inchcolm has significant restoration potential, it is possible that Brown Rat may colonise from the mainland, as it is within swimming distance. However, the risks can be reduced by adopting a robust surveillance system and incursion response plan.
- Inchcolm is one of the few remaining locations in the UK that supports Black Rat. On this basis, there may be some opposition to removing them, as was the case in the Shiants, that was also known to support Black Rat (Mcdonald & Hutchings 1997). However, responses to stakeholder consultation have so far been nearly all positive. Black Rat is not native to the UK, and is widespread throughout its native range in Asia, where populations are stable. It is also commonly encountered across the globe, where it has been introduced and has subsequently successfully colonised. Furthermore, the success of rat eradication from the Shiants provides further support in favour of their eradication from other seabird islands.
2.4. HANDA: RAT ERADICATION & BIOSECURITY
2.4. HANDA: RAT ERADICATION & BIOSECURITY
2.4.1. Site description
Site details and designations
- Handa Island is 367 hectares in area and is located ~350 m from mainland Scotland. A map of Handa, showing place names, is included as Figure A10 in the Appendix.
- Handa has high Torridonian sandstone sea-cliffs that provide tiered ledges used by a range of nesting seabird species. It is designated as an SPA and at the time of designation in 1990 supported populations of European importance for Guillemot (98,686 individuals – 9.3% of the British population and 2.9% of the North Atlantic biogeographic population) and Razorbill (16,394 individuals – 11% of the British population and 1.9% of the Alca torda islandica population). It also supports nationally important colonies of Kittiwake (10,732 pairs, 2.2% of the British population), as well as several hundred Puffins (735 AOB). The most recent counts show there are an estimated 68,524 Guillemots (individuals), 3,749 Kittiwakes (AON), 5,047 Razorbills (individuals) and 208 Puffins (individuals). These are the key species that the compensatory measure would benefit.
- As well as the key species, Handa also supports nationally important numbers of Great Skua Stercorarius skua, which numbered 66 pairs (0.8% of the GB population) at the time of designation in 1990. Since then, numbers increased to 283 pairs in 2018, with numbers in 2022 reduced to just 73 AOT. Northern Fulmar Fulmarus glacialis numbered 3,500 pairs (0.7% of the GB population) at the time of designation in 1990 but has reduced to 1,879 pairs. It also supports breeding Arctic Skua (20 AOT, SWT 2021). Other breeding species include European Shag Phalacrocorax aristotelis, Common Eider Somateria mollissima, Red-throated Diver Gavia stellata, Common Gull Larus canus, Herring Gull Larus argentatus, Great Black-backed Gull Larus marinus, Arctic Tern Sterna paradisaea, Oystercatcher Haematopus ostralegus, Ringed Plover Charadrius hiaticula and Snipe Gallinego gallinago (SWT 2021). Handa also supports a range of maritime grassland and heath vegetation.
- Seabird species that have formerly bred on Handa but were thought lost include Common Tern Sterna hirundo (last bred successfully in 2002) and Arctic Tern Sterna paradisaea (last bred successfully in 2015). However, tern chicks and fledglings (most likely Arctic) have been seen in July 2022 for the first time in 7 years. Historically Black Guillemot Cepphus grille once bred on Handa and Harvey-Brown & Buckley (1887) reported that ‘the rats had managed to dislodge them’. Also White-tailed Sea Eagles Haliaeetus albicilla once bred on Handa but have not bred since the 1800s (Harvey-Brown & Buckley 1887), although a non-breeding pair was sighted throughout the 2021 season for the first time in many years (R. Potter, SWT, pers. comm).
Site management
- Handa Island is managed by Scottish Wildlife Trust (SWT) and is part of the Scourie Estate. The island is managed by one ranger and six residential volunteers during the summer months.
- The ranger and volunteer team carry out bird counts and productivity monitoring, with Guillemot, Kittiwake, and Fulmar monitoring plots counted regularly throughout the breeding period, each year. All island counts of Guillemot, Kittiwake, and Fulmar are conducted every 5 years, whilst all island counts for terns and gulls are every 6 years. All island counts of Shag, Arctic Skua and Great Skua are carried out every 1-2 years. Other routinely recorded information includes dates of first eggs and chicks, breeding and migrant casual bird sightings, records of other notable flora and fauna sighted during the season, as well as monitoring rat activity through observations of the rat chew stations for signs of activity (see next section).
- The ranger and volunteer team also hold responsibility for ensuring that the paths around the island and the bothy itself, are maintained. They deal with regular visitor trips to the island, undertaking welcome talks and detailing conservation issues, such as ground nesting birds, fragile habitats and relevant health and safety issues. They are also responsible for producing interpretive material and for fundraising and selling souvenirs, in addition to carrying out school visits and guided walks, although these have not been taking place in recent years. However, the aspiration is to return to undertaking these activities in future years.
- Over the 2021 season Handa Island was open from the last week of March until the first week of September, with ferry crossings taking place regularly except during periods of poor weather. The island welcomed 6,661 visitors over the course of the breeding season (SWT 2021). Pre COVID-19 pandemic visitor numbers were just below 9,000 for the years 2017 – 2019 (R. Potter, SWT pers. comm.).
Invasive non-native species
- The last permanent human inhabitants of Handa left in 1848, and it is considered likely that Brown Rat Rattus norvegicus was introduced during the period between 1848-1867 when the island was farmed for sheep (Stoneman & Zonfrillo 2005). Rats were blamed for reducing numbers of Black Guillemots and for driving Atlantic Puffins ‘off the tops at Handa into more secure crevices in the face and slopes’ (Harvey-Brown & Buckley 1887). In 1904 Harvey-Brown and MacPherson stated that rats ‘simply swarm over the best ground‘(Harvey-Brown & MacPherson 1904). In 1962 Handa Island became a nature reserve, and annual wardens’ reports show that a population of rats persisted, particularly around the coast and at the bothy, the only inhabitable building on the island. As commensals, it is not surprising that their distribution is associated with human habitation. Although most cliff-nesting bird species were either stable or increasing in the mid-90s, it was felt that rats were inhibiting the expansion of Puffin, and the recolonisation of Black Guillemots. It was also felt that other ground or burrow nesting seabirds such as European Storm Petrel and Manx Shearwater might be able to colonise potentially suitable habitat were rats to be removed (Stoneman & Zonfrillo 2005). A three-year study of rats on Handa was carried out between 1994-1996 (inclusive) to better understand the potential impacts of rats on seabirds (Aragundi 1994). The work concluded that the rat population was almost exclusively limited to the coast and the bothy. The deployment of false nests showed that rats could find and decimate nests of ground nesting birds in any suitable breeding habitat. It was thought that Fulmars were not affected by rats, although it was noted that there were no ground-nesting Fulmars on Handa. It was observed that the distribution of Puffins was definitely limited by rats, with 94% of Puffins nesting on the stacks (Stoneman & Zonfrillo 2005).
- Brown rat were originally removed from Handa in March 1997 using bait poisoned with warfarin. The eradication were carried out by 12 people comprising SWT staff, volunteers and two qualified climbers. The climbers delivered and distributed bait to areas otherwise inaccessible without ropes, especially at the base of gullies and above the high tide line where rats were likely to forage. Bait was put down burrows and under rocks to avoid secondary poisoning of birds. Where there were no burrows, bait was placed under weighted fish boxes with the handles sawn out to allow rats to enter. Two tonnes of warfarin were brought to Handa. Following initial deployment, 100 kg was stored on the island for back-up baiting (Stoneman & Zonfrillo 2005). This was used in subsequent years to deal with suspected incursions.
- Following eradication efforts, a monitoring programme was devised so that staff could routinely check for further incursions. Chewsticks, consisting of lollipop sticks or wooden spatulas, were saturated in margarine oil and stuck firmly into the ground around a small bamboo marker cane. In later seasons the chew sticks were placed within plastic tunnels (thereby excluding rabbits) and sticks were replaced with wax blocks mixed with cocoa powder. These chew stations were checked monthly throughout the breeding season. Chew marks were routinely detected and there was initial confusion as to whether rats had recolonised or whether the marks were made by young rabbits. It was concluded that if rat droppings were absent then the chewing could be attributed to young rabbits. However, the regular appearance of rat droppings in 2005 (coupled with a two of suspected sightings from visitors) provided robust evidence of an incursion in that year (SWT 2005). In 2006 rat activity was also notable, with a rat observed eating a live rabbit outside the ranger’s accommodation. Tracks and droppings were also found on the boardwalk to the toilets. Rat droppings were also found in skua territories, and in August the chewsticks showed severe chewing along the southern beaches. In total rats were sighted on five occasions by visitors and rangers.
- Rat activity was also detected in 2007, with rats sighted on two occasions. Scat was found around the bothy area, and evidence of chewing was again found along the southern beaches (SWT 2007). However, by contrast in 2008 there were no rats, and Arctic terns successfully fledged 37 chicks (SWT 2008). In 2009 Biz Bell (WMIL) visited and the chewsticks were replaced with wax blocks, which are a more effective method of monitoring for rats as they show 3d imprints of tooth marks. The wax blocks were placed in boxes so that gulls and rabbits could not access them. Again, there was no evidence of rats in either 2009, 2010 or 2011 (SWT 2009, SWT 2010).
- Rats recolonised again in 2012, with signs of rat around the bothy on the warden’s return. Although six days of trapping were undertaken, only one rat was captured (SWT 2012). In 2013 there was further evidence of a growing rat population. Trapping resulted in the capture of 10 rats. The locations of the first chew marks during 2013 were found at the beach adjacent to the islet Eilean an Aigeich, suggesting that this islet and the rocks between the islet and the mainland acted as ‘stepping-stones’ for the rats. Bait stations were stationed on Eilean an Aigeich but were not checked due to lack of allocated budget.
- In 2014 SWT carried out trapping, which resulted in the capture of five rats. An additional two rats were sighted. Of these seven rats, three were sub-adults and three were adults. All were male except one (gender unknown). Tail samples were taken from two and tested for poison resistance, which came back negative. Similar trapping was carried out again in 2016, resulting in the capture of six individuals, four adults and two sub-adults, four of which were male and two of which were female.
- In 2015 data analysis was carried out using QGIS and excel to better understand abundance (measured as chew detections) and distribution of the rats. The data are included in Table 2.7, and the maps are included as Figure A9 in the Appendix. From this work it was again concluded that the rats were distributed around the coast and near the bothy.
- In 2016 it was recommended that trapping should not be undertaken except where rats became an issue, due to the labour-intensive nature of trapping. Live traps must be checked at least once daily and kill traps should be set at dusk and unset soon after dawn. This proved unworkable due to the other demands on warden time during these periods (SWT 2016). During the period between 2016 and the start of the trial of the A24 traps in 2020 trapping was only undertaken at the bothy due to reasons of hygiene, and only after a rat had been sighted. In 2016 traps were set when two rats were sighted around the bothy, both of which were trapped and humanely dispatched (one male, one female, both adults). Eight traps were periodically laid around the bothy throughout the season, but no further rats were seen or caught at this location. No non-target species were caught either. Sightings of rats were noted by wardens periodically on the Flagpole Peninsula and at Boulder Bay, however no trapping was conducted (SWT 2016).
- Over the winter of 2016/17 there was a very substantial increase in mean seasonal rat activity, which was calculated as 11 times higher than in 2015, and about six times higher compared to 2016 (Table 1 below, SWT 2017). Kill traps were set around the bothy on multiple occasions but always following a sighting, in response to the recommendations made in 2016. A total of seven rats were caught. The traps were set at dusk and unset in the morning to reduce the chance of by-catch. All people staying in the bothy were warned and shown the location of the traps when they were set. Peanut butter was used as bait and renewed when necessary and traps safely secured in place. The five rats trapped from June onwards were all found between the woodshed and the plantation. All of them were males, mostly adult. The rats were dissected, and vital statistics recorded with all rats found to be in good body condition and on one occasion feathers were confirmed in the stomach contents (SWT 2017). The step increase in rat activity suggests a discrete migration event over the winter/early spring of 2016/17.
- In 2018 rat control was only carried out around the bothy, mainly for reasons of hygiene. Kill traps were set around the bothy on multiple occasions and always following a sighting, catching 12 rats in total. Vital statistics were recorded for 11 of the 12 rats trapped, and five of them were also dissected, with all deemed to be in good body condition and none with feathers in their stomachs. One of the dissected females was pregnant with 9 foetuses (SWT 2018).
- Since 2018 rat activity appears to be gradually declining, albeit from a very high level. In 2019 the mean numbers of bait stations chewed by rats increased by 2.7%. However, trapping around the bothy (in response to sightings and primarily for reasons of sanitation) was undertaken almost every night throughout the period the wardens were in residence. A total of 24 rats were caught, and of these 22 were dissected, with all deemed to be in good body condition. One dissected rat was found with small feathers in the stomach and two hard small hard pieces that were suspected to be eggshell (SWT 2019).
- Rat activity on chew stations reduced slightly in 2020 compared to 2019 (Table 2.7). Similarly nightly trapping around the bothy area, resulted in a much smaller number of rats being captured (six only) indicating that levels of rat activity were lower. However, a decision was made in July to stop trapping around the bothy due to a blackbird being killed in one of the traps. Instead purchase of traps with longer tunnels was recommended. Three of the rats caught were dissected, but none contained bird feathers (SWT 2020). From 21 March to 6 July 2020 Handa was closed to the public due to the national COVID-19 pandemic lockdown. Trips re-opened operating from 6 July to 5 September 2020. It is possible that the slight decrease in rat activity may be due to decreased human activity, as rats are a commensal species.
- During the winter of 2020, commencing in October, the Biosecurity for LIFE started a trial of A24 traps on Handa. The project, which will run until 2023, involves trialling the use of 80 self-resetting Goodnature A24 traps to control (but not eradiacte) rats. The A24 traps can re-set up to 24 times meaning that they can be left out during the winter period when accessing the island is difficult. The traps work with a lure specifically designed for rats. The rats enter the chamber to investigate the lure triggering the trap. The body then falls out to be removed by scavengers. The traps do not use poison or other toxic substances.
- In addition, seven transects were designed across the island each containing 10 tunnels set at 50 m intervals to monitor the abundance of rats using prefabricated tracking cards. The cards would be placed in each of the 70 tunnels one day, left overnight and collected the following day. The amount of tracking cards that showed rat activity in the form of footprints or scat would be recorded. Six trail cameras were provided by Biosecurity for LIFE, along with the four already on Handa. These ten cameras were deployed to monitor traps in the vicinity of the skua colony to record potential interactions with non-target species.
Table 2.6: Percentage of chew blocks showing signs of rats (chewed/gone) across all monitoring bait stations by month, Handa Island 2015-2021.
Source: SWT 2021
- Although A24 traps have been used effectively in New Zealand, on retrieval in March 2022 it appeared as though they had not re-set. The reasons for this are currently unclear but will be investigated over the coming months (R. Potter, Reserves Manager SWT, pers. comm.). Rat monitoring in 2022 revealed relatively little rat activity early season. This is unusual (see Table 2.7) as typically more rats are detected in the early Spring when the natural food supply is less abundant (R. Potter, SWT, pers. comm.). The reasons for this are unclear, although later visits confirmed increased levels of rat activity.
- As well as rats Handa Island supports a population of European Rabbit. Historically numbers have varied: rabbits were absent in the late 1990s, but they are now so extensive that control was considered difficult and expensive (SWT 2021). However, since rabbits are not predatory their impacts on seabirds are less clear.
- In 2002 the presence of a ‘mystery mammal’ was identified through the appearance of scat. The scat was sent off for identification, and although expert consensus differed, it was generally agreed to be from European Hedgehog Erinaceus europeaus (SWT 2002). Hedgehogs, although unlikely to access sheer cliff faces, can predate the eggs of ground nesting birds such as Puffins, terns and skuas. The hedgehogs occurred again in 2003, with scat occurring in various locations. In September smaller scat was also found too, suggesting that they may have bred (SWT 2003). In 2004, it was advised by Mick Blunt of the Uist Wader Project that systematic large-scale trapping should be undertaken as a matter of urgency. In 2005 the situation become more confusing; there was a lot of scat, some of it from rats and some from hedgehogs (SWT 2005). Two volunteers spent 35 days trapping, lamping and collecting scat. Trapping was undertaken using 13 mink traps in 16 different locations and baited with cat food, mackerel or pierced eggs. A total of 735 trap nights and 22 hour 35 minutes of lamping gave no returns. However, in 2004 a hedgehog was sighted, so this and positively identifiable scat confirm that both rats and hedgehogs were present on Handa in 2005. Hedgehogs were also sighted in 2007, although there were no signs of hedgehog in 2008, 2009, 2010 or 2011. In 2012 Hedgehog scat was found again, although has not been sighted since.
- Other invasive mammals that have occurred on Handa include a stoat Mustela erminea in 2008 and in 2020, and an American Mink Neovision vision in 2021. The latter was caught using a mink trap and humanely despatched. There is also considered to be a risk of incursion by House Mouse Mus musculus, though this has never occurred. Even with dedicated effort, maintaining Handa free from invasive mammal species has been, and will continue to be, a significant challenge.
Population trends of key species
- Some background information is required on the population trends of the key species on Handa, so that and the benefits of rat eradication in 1997 can be established. This section describes population trends on Handa relative to national trends for Scotland and other relatively local sites.
- Many species have been experiencing declines associated with mass failure of sandeel prey during 2004-2008, with recent years showing some degree of recovery.
- Although Handa is located in The Minch, the population trends of many species appear to match the more severe declines associated with sites in Shetland and Orkney. It is possible that this is because of its geographical location well to the north of the other Minch sites such as Canna and the Shiants, with which it might otherwise be intuitively compared. However, otolith analysis suggests that the sandeels around Handa share a natal signature with those around Orkney, potentially reaching Handa via larval drift (Gibb et al. 2017), which would account for the severity of the declines Handa has experienced in recent decades, which have occurred despite rat eradication efforts.
Kittiwake
- JNCC SMP (Seabird Monitoring Programme) data indicates that Kittiwake numbers increased by around 24% between the late 1960s and the mid-1980s possibly due to the cessation of egg hunting (Cramp et al. 1974). However, catastrophic declines followed at least from 1986 and there are now around 50% fewer birds than in the late 1960s (JNCC 2021). Changes in the marine environment due to warmer sea temperatures, specifically the decline in the abundance of sandeels is thought to be responsible (Arnott & Ruxton 2002). Over the past decade the trend of decline has halted, and there are signs of recovery (JNCC 2021). The data from Handa, taken from counts carried out regularly at designated monitoring plots, reflects these national trends (Figure 2.5).
- However, different colonies have declined at different rates with the more northerly colonies in Scotland suffering the greatest declines in the UK. The recent 2018 seabird census survey showed that Kittiwake numbers on Handa had declined by 47% from 7,013 AON in 1999 to 3,749 in 2018. Historic data for the site shows that numbers peaked in 1977 when there were 12,500 AON. Comparison with other islands in The Minch, namely Canna, the Shiants and Mingulay and Berneray showed that other colonies have similarly declined in recent times. Kittiwakes on the Shiants declined by 46% and Kittiwakes on Mingulay and Berneray declined by 70%. The only exception locally was Canna, where Kittiwakes increased by 14%. It is of note that Black rat Rattus rattus was removed from Canna between 2005-2008, which may be the reason for the observed increases. Although rats were also removed from the Shiants, this did not happen until 2015 (the year of the JNCC survey for this site), so the benefits of rat eradication are not reflected in these figures. Although rats were removed from Handa in 1997, they had recolonised by the time of the survey in 2018. It is possible that eradication of rats from Handa may have reduced the rate of decline experienced by Kittiwake, although there is considerable variation in the rates of decline across other local sites. Comparison with the Orkney sites indicates that declines were generally more severe with declines of 47% at West Westray Cliffs, 78% at Copinsay and 84% at Marwick Head (JNCC 2021).
- Despite complete breeding failures between 2006-2008, average productivity for Kittiwake at Handa is higher than anticipated at 0.99 chicks fledged per pair, in comparison with the national average 0.83 chicks per pair for Scotland from the same time period (Figure 2.6 below, JNCC 2021). On balance it does not appear that the Kittiwakes at Handa have fared as badly as at other sites. Whether this may be attributable to biosecurity efforts is difficult to prove due to the potential influence of many other factors, but further discussion of available evidence is provided in Section 2.4.6.
Guillemot
- The Seabird 2000 Census showed that there is a general trend of increase in UK Guillemot populations since the mid-1980s to the present day. However, Scotland differs in that Guillemot numbers increased between 1969/70 and 2000, and then fell, although there is some evidence of recovery in recent years (JNCC 2021). Guillemot numbers at Handa declined from 112,767 individuals in 1998 to 54,664 in 2016, a decline of 51% (Figure 2.7 below, JNCC 2021).
- During the same period other Guillemot colonies in The Minch have also declined: Guillemots on The Shiants declined by 45% and Guillemots on Mingulay and Berneray declined by 32%. Declines in the Orkney colonies were more severe with declines of 58% at West Westray Cliffs, 65% at Marwick Head and 59% at Hoy (JNCC 2021). Although productivity was poor during the period between 2005-2008, coinciding with sandeel shortages, it has generally improved more recently, although at some sites numbers still remain low.
- For example, at Shetland productivity between 2011-13 was only 0.14 and at Orkney 0.33. Productivity at Handa was better at 0.68, in line with colonies in NW Scotland (0.58) and SE Scotland (0.70) (SWT data, JNCC 2021). Although productivity is encouraging, the scale of declines associated with sandeel failure is concerning, despite evidence of some recent recovery (Figure 2.7).
Please note that this graph shows population trends based on nest counts taken from regularly visited monitoring plots. It does not represent total Kittiwake numbers on Handa. Source: SWT 2021
Figure 2.5: Population trends of two cliff-nesting bird species on Handa Island, 1995-2021
Source: SWT 2021
Figure 2.6: Productivity of three cliff-nesting bird species on Handa Island, 1995-2021
Please note that this graph shows population trends based on nest counts taken from regularly visited monitoring plots. It does not represent total Guillemot and Razorbill numbers on Handa. Source: SWT 2021
Figure 2.7: Guillemot and Razorbill population trends, Handa Island, 1996-2021
Source: SWT 2021
Figure 2.8: Razorbill population trends based on all island counts, Handa Island, 1996-2021.
Razorbill
- Razorbill numbers in Scotland increased from 1969/70 to 2001 but declined from 2001 to 2009, remaining low between 2009-2013. They then increased up until 2017 and have sharply declined since (JNCC 2021). However, uncertainty in some of the abundance estimates means that patterns should be treated with caution. However, the abundance of Razorbills at Handa appears broadly to follow this pattern (Figure 2.7 and Figure 2.8). The last full colony count from 2019 numbered 8,207 individuals and represents a decline of 52% from 2001 (JNCC 2021). However, there is some evidence of recovery, as the previous 2014 count of 5,047 individuals represented the lowest count since records began (Figure 3.6). The Razorbill population on Handa has declined significantly in comparison with other sites in The Minch, such as Mingulay and Berneray, where the population dropped by 24% and The Shiants, where the population remained stable. The losses experienced at Handa are more in line with those from Sheltland and Orkney, that is a decline in 46% from Fair Isle and in 59% from West Westray (JNCC 2021).
- Productivity data for Razorbill has not been gathered at Handa. However, for Scotland as a whole the situation mirrors that described for Guillemot with some colonies experiencing continuing low levels of productivity (e.g. Papa Westray, and Fair Isle) whilst others have been relatively successful (e.g. North Sutor, Isle of May).
Puffin
- Estimates from the JNCC Seabird Censuses suggest that there was an increase in the UK Puffin population of 19% between Operation Seafarer (1969-70) and Seabird 2000 with an increase of 13% estimated for Scotland (JNCC 2021). The logistical issues with gathering data on Puffins mean that few colonies conduct population counts annually. However, counts at Handa have been conducted routinely, meaning that they can be compared with data from other regularly counted locations (Figure 2.9 and Figure 2.10).
- All colonies appear to have increased up until 2001 or 2002, after which numbers then declined. Monitoring on the Isle of May showed that return rates in 2007 and 2008 were particularly low. Some sites such as Sule Skerry show a recent increase, although others such as the Isle of May and Fair Isle do not. It is of note that the Handa data appears to show a significant increase in Puffin numbers following rat eradication in 1997 (Figure 2.10). Figure 2.10 also shows some recovery post 2010, followed by a decline after 2016, which is coincident with significant increases in rats (see discussion above).
- Although productivity data for Handa has not been gathered for Puffin, it has been generally low in Scotland with particularly poor years in 2007 due to food shortage and in 1998 and 2004 due to unprecedented rainfall flooding burrows (JNCC 2021). However, over the last decade it has increased, and in 2018 and 2019 reached 0.74 and 0.69 chick per pair respectively, which is similar to values from the first decade of the monitoring period (JNCC 2021).
Source: Atlantic puffin (Fratercula arctica) | JNCC - Adviser to Government on Nature Conservation https://jncc.gov.uk/our-work/atlantic-puffin-fratercula-arctica/
Figure 2.9: Number of Atlantic Puffin AOB at three colonies in Scotland, 1986-2019.
Source: Database provided by SWT
Figure 2.10: All island counts of Atlantic Puffin (individuals) at Handa Island from 1995-2020.
Population trends of other seabird species
- Other species listed as features of Handa SPA include Northern Fulmar Fulmarus glacialis, Great Skua Stercorarius skua and Arctic Skua Stercorarius parasiticus. Productivity data is collected for these species. All island counts are conducted every 1-2 years for the skuas and every 5 years for Fulmar. The skua species have been studied and ringed under the Handa Island Skua Project. Other species that are routinely counted include European Shag Phalacrocorax aristotelis, Great Black-backed Gull Larus marinus, Herring Gull Larus argentatus. All island counts for Shag are conducted every few years, but for Herring Gull and Great Black-backed Gull are conducted annually. Common Tern Sterna hirundo and Arctic Tern Sterna paradisaea are also counted annually. Common Gull has been counted on a more casual basis, since it is not listed on either the SSSI or SPA designation. Other breeding species routinely monitored include Red Throated Diver Gavia stellata.
- Other breeding species are recorded on a more casual basis. Other species that routinely breed on Handa include (but are not limited to) Common Eider Somateria mollissima, Common Shelduck Tadorna tadorna, Eurasian Oystercatcher Haemotopus ostralegus, Ringed Plover Charadrius hiaticula, Common Snipe Gallinago gallinago and Wheatear Oenanthe oenanthe.
- Other species of interest that occur on Handa, but which have not been confirmed as breeding include Black Guillemot Cepphys grylle, which is still regularly sighted on Handa. Black Guillemot is known to have bred on Handa >100 years ago, and the introduction of Brown Rat, which is first mentioned by Harvey-Brown and Buckley in 1887, is blamed for their decline (Harvey-Brown & Buckley 1887). It was suspected that European Storm Petrel Hydrobates pelagicus may breed on Handa, and mist netting was carried out on several occasions throughout the early noughties resulting in the capture of several birds, some of which had brood patches (SWT 2005). However, it could not be confirmed whether these birds were breeding on Handa as Storm Petrel is a pelagic species which may occur almost anywhere. Storm Petrel has not been sighted in recent years; however, it is a cryptic species and without specific searches for them (i.e. mist netting and or/playing calls) it is not anticipated that they would necessarily be detected.
Fulmar
- Seabird census data shows that Fulmar increased between 1969/70 and 1985-88 but has since suffered a protracted decline (JNCC 2021). At Handa Fulmars reached a peak of 3,600 AOS in 1977, levelling off in 1979 at 3,000 AOS. The recent JNCC survey in 2017 shows that following a period of stability Fulmars are now in decline, with a loss of 60% between 2000 and 2017, that is a decline from 3,550 to 1,423 AOS (Figure 2.5 above, JNCC 2021). North Rona also recorded a similar decline of 59%, although other sites in The Minch such as Rathlin, and Mingulay and Berneray did not suffer such severe declines, with numbers at these sites declining by 25% and 1.4% respectively. Productivity for Handa in 2016 and 2019 was 0.56 and 0.42, higher than the Scottish average of 0.47 and 0.39 for these years.
Skuas
- Handa supports one of the largest colonies of Great Skua in the UK and is one of the few sites where numbers had been increasing (Figure 3.9). Both skua species started nesting on Handa in the 1960s. Since 2000 Arctic Skua has been undergoing a protracted decline and have decreased from 42 AOT in 2001 to only 20 AOT in 2019, with the primary cause believed to be predation pressure from Great Skua (Jones et al. 2008). This is characterised by relatively high productivity coupled with declining numbers (Figure 2.11 and Figure 2.12). Unlike Arctic Skua, Great Skua colonies rarely experience years of breeding failure, and it is thought that this is due to their ability to switch prey, often becoming cannibalistic and also preying on the chicks and adults of other seabird species. By contrast Arctic Skua relies on stealing fish from other species (kleptoparasitism), and therefore productivity was particularly poor from 2006-2008 due to lack of prey being delivered by their host species, namely auks and Kittiwake (Perkins et al. 2018). It is of interest that Handa also supports a ‘club’ of non-breeding Great Skua at Hill Loch.
Gulls
- Small numbers of Great Black-backed Gull, Herring Gull and Common Gull breed on Handa. Great Black-backed Gull is in decline, whereas Herring Gull numbers are more stable at ~10-15 AOT. Previously Handa has supported up to 418 pairs of Herring Gull and 62 pairs of Great Black-backed Gull (in 1977 and 1970 respectively) (Figure 2.13 and Figure 2.14 below, Stoneman & Willcox 1995). The Common Gull colony numbered 31-32 pairs in the early noughties (SWT 2003, SWT 2005), having roughly doubled after rat eradication in 1997. Numbers are now very low with the latest count numbering just 9 pairs (SWT 2020). Handa previously supported very low numbers of Lesser Black-backed Gull, though breeding has not been recorded since 1989 (Stoneman & Willcox 1995).
Source: Database provided by SWT
Figure 2.11: Great and Arctic Skua AOTs at Handa Island from 1995-2019.
Source: Database provided by SWT
Figure 2.12: Great and Arctic Skua productivity at Handa Island from 2003-2017.
Source: Database provided by SWT
Figure 2.13: Great Black-backed Gull AONs on Handa Island from 1998 to 2020.
Source: Database provided by SWT
Figure 2.14: Herring Gull AONs on Handa Island from 1998 to 2020.
Terns
- Arctic Terns were last confirmed as breeding on Handa in 2015, although in 2021 Arctic Terns were observed around Port an Eilein for the majority of the season (SWT 2021). In 2022 tern chicks and fledglings (most likely Arctic) were seen for the first time in 7 years. In the early 1990s numbers of terns on Handa were small with only 1-5 pairs of Arctic Terns and fewer than 8 pairs of Common Terns (Stoneman & Willcox 1995). Numbers increased significantly following rat eradication in 1997 reaching 22 Common Tern pairs and 33 Arctic Tern pairs in 2001.
- In the same year there was also a large post breeding aggregation of 250 post-breeding Arctic Terns which arrived in mid-July and stayed for several weeks (SWT 2001). However, a series of poor years followed with various issues affecting breeding performance from sheep disturbance, unseasonal storms and high winds and seemingly unexplained nest abandonment[16].
European Shag
- In the 1970s there were reported to be as many as 400 pairs of European Shag breeding on Handa (Stoneman & Willcox 1995). In 1998 there were ~120 AON, but numbers have since fallen significantly to just 16 AON in 2020 (Figure 2.15, SWT 1998, SWT 2020). A study showed that breeding numbers in Shetland declined in a similar manner by about 87% since Seabird 2000 and it has been suggested that the majority of decline could be attributed to prolonged strong winds making foraging difficult, resulting in starvation (Wanless et al. 2018).
Source: Database provided by SWT
Figure 2.15: European Shag AONs on Handa Island from 1998 to 2020.
2.4.2. Benefits of the 1997 rat eradiaction
- Following the eradication of rats from Handa in 1997 several positive changes were recorded, the most notable being the increase in Puffin numbers. This rise is apparent in Figure 2.10 Prior to rat eradication breeding Puffins had been restricted to nesting on Great Stack. In 1999, two years after rat eradication, 20-50 AOB were counted on the main island adjacent to Great Stack. Following rat eradication in 1997 Puffin numbers on Handa increased from ~500 individuals to a maximum of 735 individuals in 2001, a 47% increase in just 5 years (Figure 2.10). It is of note that although Puffins were increasing nationally during the period between 1969/70 and 2000, on Handa they had been declining. Therefore, rat eradication had the effect of not only halting the decline but also in bringing about a very substantial population increase during this 5-year period.
- In total, seven new Puffin nesting areas were identified between 1999-2003 (2.16 below, Stoneman & Zonfrillo 2005). One of the new nesting areas was located to the south-east of Great Stack above the cliff edge and below the tourist view-point. Control of visitors was recommended by SWT wardens in 2001 and implemented successfully in 2002 but the Puffins did not return to this area again. However, this, and several other areas are still fenced off so that Puffins can potentially recolonise. Use of the additional nesting areas is noted in 2003 but these areas are not referred to again in wardens’ reports. Puffin numbers in 2004 and 2005 were notably poor (Figure 2.10). A combination prey failures, hedgehogs and rat invasion in 2005 will have contributed to current trends of decline. In some locations there is evidence of some amelioration of declines in recent years, however, this does not appear to be the case with Handa (Figure 2.10).
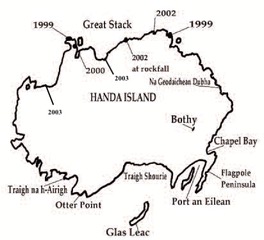
Source: Stoneman & Zonfrillo 2005 with additional nesting areas added from SWT 2003.
Figure 2.16: Map of Handa showing dates and locations of new Puffin colonies on the main island following the eradication of rats in 1997.
- Following rat eradication in 1997 Common and Arctic Terns increased, leading to some of the most successful breeding seasons ever recorded on Handa (Figure 2.17). Arctic Terns originally bred on the skerries at Port an Eilein, an islet that is cut off from the mainland at high tide. A small number were also reported to be breeding at Glas Leac, an adjacent small islet further offshore. In 1999 a new colony was established on the main island at Traigh Shourie.
- Common Gull also increased in the years following rat eradication. Initially there was a small colony of ~20 pairs at Otter Point. A new colony at Port an Eilein was observed in 1998, increasing the number of breeding pairs to ~31-32. The colony at Port an Eilein is now gone with just a very small number of pairs left at Otter Point
- A monitoring plot was established at the cliffs to investigate whether Fulmar chick survival would improve following rat eradication. Chicks were counted annually at the end of August (Figure 2.18). There was a significant rise in chicks fledged between 1999 and 2000, which could be associated with improved fledging success in the absence of rats, although this is difficult to establish due to the declines that followed.
Source: Stoneman & Zonfrillo 2005
Figure 2.17: Number of nesting Common and Arctic Terns on Handa between 1974 and 2004.
Source: SWT 2005.
- However, it was noted that following eradication there was a change in the distribution of Fulmar nests, with more nests before rat eradication on the north-west of the island, whilst after rat eradication there were more nests in the north and north-east sites. The number of nests increased in Fulmar Bay (SWT 1998), which is still in use as a nest site, although productivity from this breeding location is now notably poor (see Section 2.4.6).
- Other species were also recorded as breeding either for the first time or with increased success. These included Oystercatchers and Ringed Plovers, which were recorded fledging chicks for the first time (SWT 1998). New breeding birds for Handa included Shelduck, Redshank and Rock Pigeon, although numbers of all these were low. Sightings of Pygmy Shrew increased (SWT 1998, 1999, 2001). In 1998 extensive growth of sea rocket (Cakile maritima) and Orache (Atriplex spp.) were observed on all the islands beaches. It was thought that previously the rats consumed any sea rockets seeds. Annual growth of these species persisted until 2001, although the quantity of vegetation decreased due to grazing by sheep and the recovering rabbit population (with the latter perhaps also being a response to rat eradication).
- It was hoped that rat eradication would assist the return of Black Guillemot. Although prospecting birds were recorded in 1999-2001, this behaviour was not observed again.
- It was also hoped that eradication of rats would enable colonisation by European Storm Petrel. Although there are no records of this species breeding on Handa, it was thought that the habitat was suitable. Several searches were carried out for breeding Storm Petrels, including mist netting in 2003 and 2004. In 2004, 20% of the 96 birds netted had brood patches – one birds was a recapture from 2003. On this basis it was suggested that some birds may be breeding locally, although this was never confirmed (Stoneman & Zonfrillo 2005).
- In summary, some positive changes in seabird abundance and distribution were recorded between 1997-2003. However, the majority of species declined during the period between 2004-2008 due to large scale sandeel failures (MacDonald et al. 2019, Mitchell et al. 2020). Although rat eradiation may have alleviated the severity of the decline, the overall trend of decline was so severe that any positive effects of rat eradication are difficult to detect for many species. A similar phenomenon has occurred at Canna whereby rat eradication has had the effect of slowing the rate of decline for some species such as Guillemot and Kittiwake (Luxmoore et al. 2019). However, in recent years as prey availability seems to have ameliorated somewhat, then significant increases of both Kittiwake and Guillemot have been observed (The Seabird Group 2019).
2.4.3. Project description
- The rat eradication project at Handa is being developed in accordance with the UK Rodent Eradication Best Practice Toolkit (Thomas et al. 2017a). A brief outline of the work is included here, whilst further detail is located within the Implementation and Monitoring Plan. This includes the approach to developing a Biosecurity Plan, incorporating information on how incursion of invasive mammalian predators will be prevented, surveillance to monitor for incursion, and incursion response plans should an incursion occur.
- The difficulties in removing rats from Handa in 1997 and maintaining Handa as free from rats are significant. The number of incursions and invasions since initial rat eradication in 1997 show that maintaining Handa rat-free, or as near rat free as possible will be challenging.
- However, it is equally feasible that with dedicated effort then even islands with high reinvasion rates (i.e. greater than one rat per annum) can be kept rat free (Russell et al. 2008). Handa fulfils these criteria and more importantly is an SPA, with large nationally and internationally important colonies of breeding seabirds, and therefore leaving the rats would be unacceptable. Currently there is a significant risk of exponential increase of rats should the rats be left without further intervention.
- If greater resource could be applied to surveillance monitoring and in particular incursion response, it is feasible that Handa could be maintained as rat free in the long-term following further dedicated rat removal efforts, as it is clear the primary cause for the failure of biosecurity measure previously was lack of resource to deal with incursions. The Applicant would commit the resource required to completely eradicate rats from Handa and maintain the site as rat-free throughout the operational lifetime of the proposed development, providing the resource for additional eradication efforts as required.
- As a fundamental principle, it is necessary to understand where the rats are coming from so that they can be controlled. Although the first records of rats re-occurring on Handa in 2012 were near the stepping-stone islets, it is unclear whether the rats are coming from Tarbet or elsewhere along the mainland. Identifying and controlling the source of the rats and establishing a rat free buffer area along the mainland would reduce future incursion risk.
- The relevant stakeholders are supportive of rat eradication, although continuing engagement, and engagement with a wider range of interested parties (e.g. the local fish farm) will also be undertaken to ensure support for the work is sustained over time.
- It is anticipated that the eradication phase will be conducted during the winter period by an eradication specialist in collaboration with SWT, as detailed within the Implementation and Monitoring Plan.
- The rodent eradication will be funded by the Applicant. The Applicant will also supply resource to maintain biosecurity at Handa throughout the operational lifetime of the Proposed Development. The resource would cover monitoring of bait stations, and any trapping required in future should incursions occur.
2.4.4. Conservation targets
- The benefits of rat eradication are likely to be influenced by a range of factors. These are listed in section 2.3.5. However, comparison of seabird numbers, distribution and productivity data from Handa gathered in years with and without rats provides some indication of the benefit that may be achieved by a second rat eradication programme.
- Since the rats have returned to Handa, the Puffins are again restricted to breeding on Great Stack, a huge 120 m tower of Torridonian standstone, which matches the height of the adjacent cliffs and which also supports a similar number of nesting ledges. The stack is almost impossible to climb, and it is believed that it has always remained free of rats. This would seem to be a reasonable assumption.
- There are other smaller stacks including Needle Stack and a couple of ‘stacans’, or little stacks, which may also be free of rats. However, all the key species breed on Great Stack and at other locations on the main island. Therefore, comparison of productivity data (when available) from Great Stack with that obtained from mainland breeding locations enables some assessment of the level of impact that rats may be having. However, it is acknowledged that there are other factors that could also account for the difference between Great Stack and the main island such as habitat quality and human disturbance. The similarity in habitat between the cliffs adjacent to Great Stack and Great Stack itself is notable, as both are structurally identical, formed from the same Torridonian sandstone and both with the natural ledge formations that form ideal nesting habitat for many seabird species. However, other cliffs and breeding locations around Handa are not necessarily comparable in terms of habitat. It is also not possible to separate the impact of human disturbance: Great Stack is completely inaccessible, whereas other breeding locations are at varying distances from the path.
- In all instances the predicted benefits are viewed as precautionary in the sense that they do not incorporate how additional birds supplied annually will contribute to the growth of the colonies over the project lifespan.
Puffin
- Puffin numbers increased from 472 Puffins in 1996 to 735 Puffins in 1997 (Figure 2.10). This equates to a 56% increase and a gain of ~44 Puffins per year. However, this rough measure of benefit does not adequately describe the effect that an increased number of adults, together with increased productivity would have on promoting colony growth. However, understanding this process fully would be complex as other factors such as the allee effect and density dependence would need to be incorporated. Equally it is also acknowledged that the initial 1997 rat eradication took place when Puffin numbers were increasing nationally, although the step increase following rat eradication is obvious (see Figure 2.10). It is also important to appreciate that Puffin started to colonise the main island immediately after rat eradication, strongly indicating that rats were restricting Puffin to Great Stack.
- It is considered realistic that this level of increase could be achieved again, along with recolonisation of the main island, especially if areas are protected from human disturbance in advance of the eradication. It is certainly feasible that the colony could, in time, return to the numbers observed in 2004 (735 individuals). Indeed, if conditions were favourable, Puffin could exceed these numbers. An NVC survey carried out in 1997 suggested that there is ~20 ha of maritime cliff habitat (excluding ledges) that would be available to Puffin. Assuming a burrow density of 0.5 burrows per m2, based on what has been observed at St Kilda (Harris & Rothery 1988) it is feasible in terms of habitat alone that Handa could support >100,000 Puffin burrows. Although there are many reasons why this may not be achieved in practice, habitat availability is clearly not a limiting factor with regards to achieving conservation targets.
- The benefits of rodent eradication for Puffin as achieved at other islands are discussed fully in Section 2.3.5. At present Puffins on Handa are restricted to Great Stack, which is a breeding location also favoured by other species, presumably because of the lack of rats (and possibly human disturbance). It is possible that productivity could improve if other breeding sites were made available through the eradication of rats from the main island. If nothing is done to improve conditions for Puffin and the current pattern of decline continues, which over the past 20 years approximately averages around 25% per year, then it is considered likely that Puffin will be lost from Handa as a breeding species by around 2030.
Guillemot
- Guillemot numbers did not increase following rat eradication in 1997 (Figure 2.7). Although it is likely that there will be some benefit to Guillemots from rat eradication, it is not expected that the effects would be as marked as for Puffin and Razorbill. Guillemots nest on cliff faces, laying a single egg on a narrow ledge rather than building a nest. In some locations these ledges may be accessible to rats, as rats will climb cliff faces, but in many instances they may not be.
- Guillemots breed in roughly 11 different locations around Handa, and their distribution has not changed significantly since rat eradication in 1997 until recent years. Figure 2.19 shows an increase in the number of Guillemots nesting on Great Stack from 2018 onwards, which is coincident with the sudden increase in rat numbers. Guillemots appear to have relocated from G4 (Am Bonair) to Great Stack, which is located nearby offshore. It is of note that Am Bonair is one of the locations that Puffins recolonised following rat eradication in 1997 (see Section 2.4.2), suggesting that rats may be able to access nests in this location. Maps of rat activity (see Figure A9 in Appendix) show that rat detections increased in this location in 2017 perhaps resulting in a lower number of nests in this area in 2018. In summary, it appears that at least some Guillemots may be impacted by rats and are being displaced from Am Bonair on the mainland to Great Stack. Numbers of Guillemot nests on Great Stack have increased from 361 AON in 2018 to 436 AON in 2021 (an increase of 21%).
- Productivity monitoring for Guillemot has been carried out since 1997 with monitoring plots located at Poll Ghlup Geodh, a sheer sinkhole formed from a collapsed sea cave behind the cliff edge, and on the Great Stack. Therefore, it is possible to compare productivity on the mainland against productivity on Great Stack after the first rat eradication, and more recently now rats have returned. However, there are caveats: Poll Ghlup Geodh (G9 on Figure 2.19) may well be difficult for rats to access due to the sheer nature of the cliffs. It is also impossible to control for the influence of human disturbance. Poll Ghlup Geodh lies on the footpath, although people cannot closely approach the birds.
- During the rat free years, Poll Ghlup appeared on most occasions to be the most successful breeding location in terms of productivity (Table 2.8). However, during the recent rat invasion productivity has generally been higher on Great Stack. It is of note that this change occurred between 2016 and 2017, which ties in with the step increase in rat activity shown in Table 3.10. The difference in productivity between Poll Ghlup and Great Stack without rats was 0.07, but with rats productivity at Great Stack was 0.01 higher. Assuming that productivity at Great Stack and Poll Ghlup are roughly equal when rats are present, then it could be estimated that rats decrease productivity by ~0.05. As mentioned previously there may be other factors that could also affect productivity, which it is not possible to separate further.
Figure 2.19: Number of Guillemots nesting in different areas around Handa Island from 2015-2021, during the period of rat invasion.
- The impacts of rats on Guillemot productivity may differ at other nesting locations, such as Am Bonair, based on their accessibility to rats. This is an area which would require further study to progress understanding about the impacts of rats on Guillemots across all the island nesting locations.
- However, even a small difference in productivity can result in an increase in numbers if applied to a colony the size of Handa. For example, based on the latest SWT count of 68,524 Guillemots, then even an increase in productivity of 0.05 would constitute an additional 1,713 additional fledglings per year. Using the mean age specific survival rates from Horswill & Robinson (2015), and assuming a mean age of breeding of 6 years (Harris et al. 2016) then this increase in fledglings would translate to the supply of 577 additional adults per year.
- Since historically Guillemot numbers have been much higher, numbering 98,686 individuals in 1990, then it is clear that Handa does support enough suitable habitat to accommodate the potential increases that rat eradication and implementation of biosecurity measures could deliver over the operational lifetime of the Proposed Development.
Source: Compiled from wardens reports SWT 1997-2002 inclusive.
Razorbill
- Razorbill numbers on Handa have declined significantly, although there are some indications of recovery (see section 2.4.1). Although specific monitoring plots are counted annually (in the same locations as the Guillemot monitoring counts), all island counts are only conducted every 5 years.
- Comparison of all island Razorbill counts between 1997 (the year of rat eradication) and 2001 show that Razorbills increased from 15,573 to 17,042, an increase of 1,469 individuals (or 9%). Although this is in line with national trends (see Section 2.4.2) there were notable increases in use of some specific areas. These included An Carn Dubh boulders where numbers increased from 394 to 704. It seems likely that the presence of rats was decreasing habitat quality. By 2019, numbers of birds in this area had declined to only 195 Razorbills. Habitats such as boulder fields have been recolonised in other islands where rats have been removed such as Ailsa Craig.
- Numbers of Razorbills nesting on the main island cliffs opposite the Great Stack also increased from 609 individuals in 1997 to 1,127 in 2001 (an 85% increase). It is of note that Puffins also recolonised these cliffs during this period, indicating that rats may be able to access at least some of the nests on these cliff faces. In 2019 there were only 555 birds in this area, in line with numbers from 1997.
- Similarly, numbers of Razorbills increased at Geodh Great Stack from 499 birds in 1997 to 757 birds in 2001 (an increase of 52%). In 2019 this count area was combined with counts from Great Stack where numbers had significantly declined (see above).
- Numbers of Razorbills at Goedh Dearg increased from 740 in 1997 to 1182 in 2001 (an increase of 60%) suggesting that this usage of this area increased following rat eradication. Recently numbers have significantly declined in this nesting location to just 490 nests in 2019 (a decline of 141%), again suggesting that rats may be able to access this area.
- Although the Razorbill population on Handa did not increase following rat eradication, it seems likely that rats were preventing Razorbills from using some of the available (and otherwise suitable) habitat on the main island including An Carn Dubh boulders, the cliffs opposite Great Stack and Geodh Great Stack. It is likely that use of these areas would increase should rats be removed. However, annual all island counts and productivity monitoring of different areas of representative habitat would be required to quantify the benefits of rat eradication.
- Since Razorbills nest either on lower cliff ledges or among boulders at the bottom of cliffs, then significant improvements to breeding success following rodent eradication are anticipated. Indeed, there is evidence of increases in Razorbill numbers at 5 islands where they have been monitored following rodent eradiation, namely Canna, Lundy, Ramsey, the Shiants and Ailsa Craig (Thomas et al. 2017a, Brooker et al. 2018, RSPB News from the Rock 2021). However, Razorbill has been increasing nationally and so caution is required in interpreting this information.
- For example, rodents were removed from the Shiants in 2015, and by 2018 productivity for Razorbill increased from 0.72 to 0.79. Assuming a similar rise in productivity of 0.07 for Handa and based on the latest SWT count of 5047 individuals, then an increase in productivity of 0.07 would result in an additional 353 fledged chicks per year. Using the mean age specific survival rates from Horswill & Robinson (2015), and assuming a mean age of breeding of 4 years (Lavers et al. 2008) then this increase in fledglings would translate to the supply of 160 additional adults per year.
- Since historically Razorbill numbers have been much higher, numbering 16,394 individuals in 1990, then it is clear that Handa does support enough suitable habitat to accommodate the potential increases that rat eradication and implementation of biosecurity measures could deliver over the operational lifetime of the Proposed Development.
Kittiwake
- Kittiwake productivity monitoring is carried out at eleven plots annually. All island Kittiwake counts are carried out every five years. The timing of the counts (1999 and in 2005) is such that the impacts of sandeel failure from 2004 onwards dominate the dataset, which broadly reflects national trends (see Figure 3.3). However, it is possible to compare productivity between the monitoring plots on the main island and on Great Stack both from 1997-2002 representing the period after rat eradication and from 2015-2021 when rats had returned. This was done using a Wilcoxon signed rank test which showed that birds nesting on Great Stack were significantly more successful than those nesting on the main island both when rats were present on Handa (2015-2021)[17] and when they were absent (1997-2002)[18]. This suggests that the Great Stack represents superior habitat for Kittiwake over both time periods (Table 2.9).
Table 2.8. Productivity (number of chicks fledged per nest) of Kittiwake nesting on Great Stack and the main island during periods when rats were absent (1997-2001) and present (2015-2019)
Source: Compiled from wardens reports SWT 1997-2002 inclusive.
- However, the size of the difference is greater during the period between 2015-2021 (average difference =0.30) compared with 1997-2002 (average difference =0.19). This suggests there is an increase in average difference in productivity between the main island and Great Stack of 0.11. It is suggested that this could be attributed to rats, although may also be influenced by other factors (e.g. human disturbance) which cannot be separated out.
- Based on the last all island Kittiwake count numbering 3,749 AON from 2018, and assuming that 31% of Kittiwakes nest on Great Stack and 61% on the mainland (as in 2021[19]) then the 2,287 pairs nesting on the mainland could increase their productivity by 0.11, resulting in an additional 251 birds fledging per year. However, this data is based on comparison of two sites, and comparison of productivity between other nesting sites around Handa Island is required to generate a fuller understanding of the impacts of rats on Kittiwakes. Using the mean age specific survival rates from Horswill & Robinson (2015), and assuming a mean age of breeding of 4 years (Coulson 2011) then this increase in fledglings would translate to the supply of 124 additional adults per year.
- Since historically Kittiwake numbers have been much higher, numbering 125,000 individuals in 1990, Handa clearly does support enough suitable habitat to accommodate the potential increases that rat eradication and implementation and biosecurity measures could deliver.
Summary
- The benefits that may be delivered by rat eradication at Handa for the key species are summarised in Table 2.10. However, these numbers are potentially an underestimate of the benefit that may actually be delivered. As more adults are added to the population, the number of additional fledged chicks (and therefore adults) increases over time. The impact of the regular addition of these adults and their contribution to colony productivity is not recognized by these calculations. It is also considered likely that once rat free the numbers of adults that choose to nest on Handa would increase due to redistribution of birds from other local colonies (as appeared to occur after rat eradication in 1997 – see Figure 2.10). The contribution of these birds to the growth of the Handa colony is also not captured by this approach.
- Conservation targets for Handa have been set by multiplying the number of additional adult birds per year that would be generated by rat removal (as calculated above) by the 35 year project lifespan, and then adding to this the number of birds that are currently present. In all species except Puffin, historic maximum counts exceed conservation targets, demonstrating that Handa has the habitat to support these increases. The only exception to this is Puffin. However, as mentioned above Handa has the habitat to support >100,000 Puffin burrows and therefore habitat availability is not considered to be a limiting factor. Although the conservation target for Puffin exceeds historic maximum counts, it is considered to be realistic in view of the success of other rat eradication projects for this species (see Section 2.3.5).
2.4.5. Benefits to other species
- It is considered feasible that the eradication of rats will facilitate the return of Arctic Tern to Handa as a regularly breeding species once again. It is also possible that Common Tern could breed again too. Arctic Tern numbers increased significantly following rat eradication in 1997 (see Figure 2.17). The recolonisation of Feno Island (Terceira, Azores) by Common Terns and Roseate Terns was achieved following rat eradication (Amaral et al. 2010). Since terns are ground-nesting then the impacts of rats will be significant and improvement following eradication would be anticipated based on previous experiences from both Handa and elsewhere.
- It is unclear to what extent rat eradication will improve the breeding success of Fulmar, although some benefit would be anticipated. Rat eradication has resulted in improved breeding success of Fulmar at Lundy (Brooker et al. 2018) and Ailsa Craig[20], although not at Canna where the nesting area is not considered to be very accessible to rats, and where Fulmar numbers have continued to decline despite improvements in many other species (Luxmoore et al. 2019).
- At Handa there was some evidence of increased chicks fledging from the cliff top nest sites in 2000, although (like many of the initial improvements observed on Handa) this was not sustained. On Handa Fulmars nest in four locations: Great Stack, Little Stack, Puffin Bay and Fulmar Bay. Following rat eradication in 1997 the distribution of nests shifted, and Fulmar numbers increased in Fulmar Bay. Fulmar Bay is now the smallest nest site with 10 nests in 2021 compared to 41 on Little Stack, 52 on Great Stack and 49 in Puffin Bay. Comparison of productivity between these sites suggests that Fulmar Bay may well be impacted by rats (see Figure 2.20), with productivity reduced by about 0.20 relative to the other sites.
Figure 2.20: Mean productivity of Fulmars nesting in different locations on Handa Island. Data from 2015-2021 (excluding 2020 when data could not be gathered).
- It is not anticipated that Puffin Bay is impacted by rats, and this perception is reinforced by historic data, which shows a continuation of the trend of decline with no improvement following rat eradication in 1997 (Figure 3.21).
Figure 2.21: Fulmar AOS counts at Puffin Bay, Handa Island (1983-2005).
- Although this is not a species that has been studied in relation to rat eradication from islands, it is anticipated that there would be a benefit to Red-throated Diver. There are usually several pairs that breed on Handa (5 pairs in 2021 – SWT 2021).
- Eradication of rats has been of benefit to Shag on Ailsa Craig where they colonised following rat eradication and Canna, where numbers increased particularly in boulder field habitats (Luxmoore & Bell 2019). On Lundy, where Shag had been in decline, the eradication of rats served to halt the decline resulting in a stabilisation of numbers (Brooker et al. 2018). There is some evidence of this type of pattern on Handa following rat eradication in 1997 (Figure 3.13) before numbers declined sharply after 2001. On this basis it is anticipated that there would be a benefit to Shag from removing rats, as numbers are now very low. During the all island Shag count conducted in 2020 there were only 16 AON left, which represents a 50% decline since 2015. If rat eradication could halt this decline, it could prevent this species from being lost from Handa.
- Common Gull benefitted previously from rat eradication in 1997 and it is considered likely increased numbers could again be achieved. Numbers are currently very low (9 pairs), and like Shag, could be lost from Handa as a breeding species if the trend of decline continues. Some benefit to Herring Gull and Great Black-backed Gull is also considered possible, although numbers of both on Handa are relatively low.
- It is unclear whether rats are impacting on either Great or Arctic Skua, and this is an area that requires further study.
- It is anticipated that rat eradication would bring obvious benefit to ground nesting species present on Handa such as Oystercatcher and Ringed Plover. Benefits to these species were recorded after rat eradication in 1997. Dunlin and Redshank were also observed breeding for the first time on Handa after 1997, although now they no longer do. It is possible that these and/or other shorebirds could colonise and breed if Handa were free of rats again. Benefits post 1997 were also recorded for Eider and Shelduck.
- Other breeding species that occur on Handa considered likely to benefit from rat eradication include (but are not limited to) Cormorant, Grey Heron, Red Grouse, Snipe, Rock Dove, Skylark, Meadow Pipit, Pied Wagtail, Dunnock, Wren, Blackbirds, Song Thrush, Robin, Stonechat, Wheatear, Goldfinch and Willow Warbler.
- Since rat eradication did not previously result in the recolonisation of Storm Petrel, Black Guillemot or Manx Shearwater as had been speculated, then it is not anticipated that this would occur. However, it is difficult to predict what may or may not occur and the continued sighting of Black Guillemot does offer some hope.
- Following rat eradication in 1997 there were increased sightings of Pygmy Shrew. Since rats are a predator, then increased Pygmy Shew numbers would be anticipated. This is a phenomenon that has also occurred on Ailsa Craig and Lundy (Thomas et al. 2017a).
- It is also considered likely that rat eradication would benefit invertebrate populations on Handa. There are several notable butterfly and moth species, and it is considered possible that reduced predation of caterpillars by rats could be beneficial in increasing abundance and species richness of lepidoptera.
- It is also likely that Rabbits will increase following rat eradication as has happened on the Shiants, where some control of the increasing rabbit population has been required. After the 1997 rat eradication Rabbit numbers were low (possibly they were vulnerable to warfarin poisoning) but increased rapidly in the absence of rats. This then resulted in grazing of Sea Rocket beds, which had increased rapidly following rat eradication. The possibility of controlling rabbits should be considered at the project design stage to avoid similar scenarios reoccurring. It may not be necessary to control Rabbits if there are no apparent detrimental impacts, but it is necessary to be able respond rapidly to changing situations as appropriate.
2.4.6. Timescale & mechanism for delivery
- Full details of how the project will be delivered are included within the Monitoring and Implementation Plan, and only a brief summary is included here.
- It is anticipated that the initial rat eradication phase will be undertaken by an eradication specialist during the winter months, with surveillance and seabird monitoring conducted by a qualified contractor, who would also be responsible for implementing incursion response plans should an incursion occur.
- Since stakeholders are positive, it is not anticipated that there are any significant barriers to implementation, which could be undertaken relatively rapidly (although the eradication phase itself would need to be undertaken during the winter months).
2.4.7. Additionality & uncertainty
Additionality
- Since Handa Island is an SPA where biosecurity work is already underway then the issue of additionality does need to be addressed. This issue was raised by the RSPB following the final stakeholder consultation meeting on 22nd September[21]. They suggested that the removal of rats from Handa would not be an appropriate compensatory measure as commitments to biosecurity should already be incorporated within the current SPA management objectives. Whilst Handa does have management objectives related to maintaining biosecurity, these are to monitor for the presence of Brown Rat and control if necessary. They do not mention the eradication of Brown Rat, implying that their aim is to minimize rat numbers only.
- The recent invasion of rats in 2012 clearly demonstrates that the level of resource SWT are currently able to dedicate to biosecurity is not sufficient to control rat numbers. This is demonstrable through reference to the wardens reports (SWT 2016-2020), which show that staff were aware of the developing problems with rats, but were unable to dedicate staff time to setting and checking the large number of traps required to control what was clearly a rapidly increasing rat population.
- The lack of financial resource to charter the ferry to check bait stations deployed on one of the stepping-stone islets was also a problem (SWT 2012). Although the bait stations were installed in 2012 as it was understood that the islets were highly likely to be the route of recolonisation, there was no resource to carry out checks of these bait stations.
- Both EC and national guidance on compensatory measures indicates that measures that are accepted as ‘normal’ practice should not constitute compensation. ‘Normal’ practice is defined as being within the bounds of everyday financial and political realities. However, if normal practice is failing, and the measures suggested are additional (i.e. over and above what can reasonably be expected to happen in the absence of the project), then the work can be classified as appropriate compensation.
- Although SPAs would be expected to maintain biosecurity by writing a biosecurity plan to minimise the chance of incursion and to carry out surveillance monitoring, it is argued that maintaining biosecurity at an island such as Handa will always require resource over and above what may typically be required at other islands. This is due to a combination of factors: its proximity to the mainland, the presence of several ‘stepping-stone’ islets between Handa and the mainland, its large size, the number of visitors, and the availability of ample habitat and natural food resources for rodents. For these reasons, coupled with the high conservation value of the site for seabirds, significant additional effort will always be required to control rats at this site.
- The range of potential measures under consideration for Handa are by any definition over and above the current management objectives for the site, which at present offer no commitment to rat eradication in spite of the obvious need for rats to be removed both due the high conservation value of the seabird colonies and the known negative impacts of rats on seabird islands.
- SWT currently employ one paid warden on a seasonal basis to manage the reserve at Handa. The warden is assisted by five volunteers. The warden and volunteers run the reserve, carry out seabird counts and productivity monitoring, meet visitors and ensure they keep to the paths (several ferry trips a day), man a small souvenir shop, and carry out school visits, repair work and general maintenance of the bothy area.
- There is therefore currently little resource to dedicate to biosecurity. On occasions when rats have reoccurred the there is no resource to currently fund further systematic rodent control or eradication efforts. This has ultimately led to reinvasion of rats and a return to high levels of rat activity
- The Applicant would supply the relevant resource to deal with all eradication, monitoring and elements of biosecurity. This would include a resource to undertake rat eradication work if further incursions of rat and other invasive mammalian predators occur. Since rat chew stations are currently monitored, there is a small element of additionality with regards to this single element. However, at present surveillance visits are carried out monthly during the breeding period. A dedicated biosecurity resource would facilitate improved surveillance with increased frequently of visits including during the winter period. Other tasks such as looking for scat, carrying out lamping for hedgehogs, and trapping for rats and routine checking of any traps that are set could also be carried out routinely with additional resource.
- Funding would also be supplied to coordinate a dedicated rat eradication in the event of future invasion. This is of particular importance, since failure to fund incursion response can be viewed as the primary cause of failure previously. The funding would also facilitate the sustained stakeholder engagement required for the biosecurity work to be successful, enabling SWT to engage with a wider range of stakeholders than has been undertaken previously.
Uncertainty
- There is reasonable certainty that rats can be eradicated from Handa, as this has previously been achieved successfully. However, there is uncertainty around whether the current A24 traps are effective. However, alternative methods of rat eradication will be pursued.
- Future incursions and potential reinvasions are a risk, and indeed would be anticipated at a site such as Handa. However, the additional resource provided by the project would enable both continuous surveillance and rapid response to such events.
- There is some uncertainty regarding the conservation targets set in Section 2.4.5. Quantifying the benefit rat eradication is difficult; comparison of counts before and after rat eradication may be of limited use as it is impossible to separate out the influence of other factors, such as mass prey failures, which may have a significant impact on the dataset. This point applies to Puffin only as productivity data has been used to predict benefits for all other species.
- Applying data from one site to make predictions about another is problematic as the islands may be subject to different pressures and may therefore respond differently to rodent eradication. This point applies only to Razorbill, where data from the Shiants has been used to inform predictions for Handa.
- However, predictions for Kittiwake and Guillemot are probably as accurate as can reasonably be expected as they are based on comparisons with rat and rat free nesting areas and use long-term site-specific productivity datasets. However, in all instances the predicted benefits are viewed as precautionary in the sense that they do not incorporate how the additional birds supplied annually will contribute to the growth of the colonies over the project lifespan.
3. TIER I: SAFEGUARDING KITTIWAKES AT DUNBAR
3. TIER I: SAFEGUARDING KITTIWAKES AT DUNBAR
3.1. Background
3.1. Background
- Dunbar Castle, harbour and the adjacent coast supports a considerable number of breeding seabirds, with >800 pairs of Kittiwake and smaller numbers of Northern Fulmar (16 pairs in 2020), European Shag (16 pairs in 2020) and Herring Gull (15 pairs in 2020). The compensation at Dunbar would therefore be of benefit to Kittiwake only and not the other key species.
- Although numbers are low compared to the larger colonies, such as St Abbs and the Isle of May, Dunbar supports more Kittiwakes than Coquet and more than any of the Forth Islands, with the exception of the Isle of May (Figure 3.1) making it a significant local colony.
- Funding a warden for the Kittiwake colony at Dunbar Castle (a non-designated site) was recommended during stakeholder consultation to improve the numbers of adults nesting at Dunbar and their breeding success. Improving breeding success at Dunbar would strengthen Kittiwake populations within the Forth Islands and Farne Islands SPAs. Ringing of Kittiwakes at Dunbar between 1993 and 2007 resulted in the capture of 16 ringed birds from other sites, proving connectivity with the Isle of May (10 birds), Inchkeith (4 birds) and the Farne Islands (2 birds) (Coleman et al. 2011)[22].
- Problems with the site identified during stakeholder consultation included human disturbance, and discarded fishing nets left around the harbour, which then become incorporated in Kittiwake nests leading to risks of entanglement/ingestion (see Section 3.3 below). It was felt that a warden would be able to liaise with both the public and the fishermen to resolve these issues. Further consultation with East Lothian Council (ELC) and the Dunbar Harbour Trust (DHT) confirmed this assessment of the situation, and the benefits of having a ‘Kittiwake warden’ were unanimously agreed.
- Following the Applicant’s Compensation Consultation meeting 30 March 2022, concerns were raised by NatureScot regarding the uncertainty and time that it would take to research and implement the measures suggested for Kittiwake at Dunbar. It was suggested that poor prey availability could be the driver for the decline at Dunbar and that wardening may not be an effective method to increase productivity. The need for habitat enhancement was questioned on the basis that there is spare nest capacity with possibly 300-500 spaces already available. The unquantified nature of the disturbance impacts, and the anecdotal nature of the evidence was also raised as being of concern[23].
- Further work regarding the disturbance issues at Dunbar and evidence regarding the prey situation from Searle et al. 2022 (in prep.) were presented at the Compensation Consultation meeting on 8th June 2022 and are discussed in full within this report. Following this additional work there was a consensus from NatureScot, Marine Scotland Science and RSPB that human disturbance is an issue at Dunbar, and that reducing disturbance ‘would likely improve productivity of the Dunbar Kittiwake colony’[24].
- The focus of comments related to: i) how the benefits of wardening would be assessed and quantified, ii) to what extent the wardening would provide ‘significant measurable benefits’, and iii) the need for evidence relating to the various types of human disturbance and which may be the most damaging for the Kittiwakes. The anecdotal nature of the evidence was again raised, as was the requirement to account for the adverse impact of the Proposed Development on the Dunbar Kittiwake colony as a component of this assessment[25].
- A brief summary of how the benefits of wardening could be measured, quantified and assessed, is included in Section 3.3 and further details are included in the Implementation and Monitoring Plan. Further study of the various types of human disturbance and their impacts on the Kittiwakes would need to be carried out.
- It is not possible to define to what extent this project will improve colony productivity in advance of the work being undertaken. However, Section 3.3 sets out a series of hypotheses and how these could be tested. As a starting premise, the data presented in Searle et al. 2022 (in prep.) suggests that prey is not a limiting factor, therefore implying that colony-based issues are driving the decline in the Dunbar Kittiwake colony. In essence this project allocates resource to systematically and scientifically investigate and tackle the various factors that may be impacting negatively on the birds to improve productivity. For this reason, monitoring and adaptive management is recommended so that it can be agreed that the resources are being directed appropriately and that the project is delivering measurable benefits. It is anticipated that the data from the project could be reviewed annually. This approach has the benefit of allowing the project to evolve in accordance with the needs of the site, which are likely to change over time.
- As the site is neither a SSSI nor an SPA it does not currently have a dedicated warden, although the birds are counted annually by the East Lothian Countryside Ranger Service (ELCRS), who have also monitored colony productivity in the past. Kittiwakes from Dunbar are also ringed by a local group. Since there is no resource to gather any other data from Dunbar, the information presented within this report is to a certain extent anecdotal, although many observations are from the local ELC Warden with >20 years of experience working within this Dunbar Castle area and are considered (in the absence of any other formally gathered data) to represent the best available information.
3.2. Site description
3.2. Site description
3.2.1. History of the Dunbar Kittiwake Colony
- The Kittiwake colony has a well-documented history due to its accessibility. In 1934, 3 nests were recorded. The colony increased between 1934 until the late 1950s reaching 201 nests in 1959 (Coulson 1963, 1983). It then underwent a period of decline from 1959–1976, with 189 nests in 1976 (Coleman et al. 2011). From 1976–2000 the colony continued to increase. Prior to the 1980s, the Kittiwakes nested in the Granary, although they abandoned that nesting site when it was renovated. It is unlikely that they will nest there again, as it is now occupied.
- During the 1980s the Kittiwakes expanded to occupy nesting areas around the North and South Harbour entrances and the Magazine (a former weapons store). In 1995 a large quantity of masonry from the Castle fell into the sea, and the Castle was closed to visitors on health and safety grounds.
- It seems likely that at this point a reasonable proportion of the Kittiwakes nesting on the main colony relocated to the Inner Castle, which was no longer accessible to people (see Figure 3.4). Use of the cliffs adjacent to the Leisure Pool also started around this period, and the Kittiwakes also started to nest on the various rock outcrops, which were formed by the collapse of the Castle into the sea. The nesting area on the leisure cliffs is of specific interest as the number of birds in this location have been gradually increasing, numbering 152 nests in 2021. This area is not accessible to people (Table 3.1, Figure 3.4).
- The Kittiwakes now nest in a variety of locations around the main Castle, ruins, harbour and surrounding cliffs (Figure 3.1, Table 3.1). However, they do not nest on any residential buildings as is often the case in urban locations.
- The number of Kittiwake nests on the main Castle has been steadily declining since the late 1990s, although total numbers of Kittiwake nests at Dunbar continued to increase until the early noughties reaching a peak of 1,191 nests in 2000. However, it is the sustained decline in nests on the main Castle that appears to be the driving the general trend of decline at this site (Figure 3.3 and Figure 3.4). The most recent count from 2020 recorded 808 nests, with only 153 nests on the main Castle (Figure 3.3, Table 3.1). Since the habitat itself is unlikely to have changed appreciably, and the main Castle remains in many ways superior to some of the other areas used, such as the Sea wall which is regularly flooded out and the outcrops which are lower and very exposed, it is speculated that the decline may be due to increased human disturbance and could potentially be rectified.
3.2.2. Stakeholders & ownership
- Dunbar Castle was formerly owned by ELC, who still have responsibility for monitoring Kittiwake numbers. Ownership of the castle itself was transferred in 2004 to the Dunbar Harbour Trust (DHT), a charity set up by a number of harbour users with the aim of improving the harbour facilities for the professional and leisure users. The Board’s mission statement is to ‘run a safe, efficient and welcoming harbour that caters for the needs of all the harbour users, visitors and the local community as well as the environment’.
- One of the strategic objectives identified by the Board is improvement of the environment within the harbour. The Castle itself is a Scheduled Monument, and the whole of Dunbar Harbour is a historic conservation area. Although the Castle has played an important role in Scotland’s history, it is in considerable disrepair and there are no plans to restore or conserve it, although work to prevent further collapse is occasionally undertaken.
- Dunbar Harbour itself is small with 28 fishing vessels and around 60 members. It is also popular with leisure users, and every leisure mooring is currently taken. The fishing vessels are 6–15 m in length and generally fish locally for Norway Lobster (Langoustine) Nephrops norvegicus and crabs. The Harbour is inaccessible to larger vessels as the entrance is difficult to navigate and has problems with surge.
3.2.3. Number and location of Kittiwake nests
- The number and locations of Kittiwake nests are shown in Figure 3.2, with Table 3.1 showing how the distribution of nests has changed over time. Changes in the number of birds nesting in each of sub-sites is plotted in Figures 3.3 and 3.4.
- The decline in nests on the main Castle described above is mirrored by similar declines on the north Harbour entrance and the south Harbour entrance, with declines in all three areas starting during the mid to late 90s (Figure 3.4). In the mid-90s, there were ~250 nests on the south Harbour entrance and >100 nests on the north Harbour entrance. By 2020 this had declined to 132 nests on the south Harbour entrance and 20 on the north Harbour entrance.
- The main Castle, the north Harbour entrance and the south Harbour entrance can all be accessed via a footpath. The path that runs along the main Castle colony and the green in front of the steps are the key areas where there is greatest overlap between the public and the Kittiwakes. At present people, dogs and children (in unlimited numbers) are able to walk along the narrow path that directly abuts the outer wall of the main Castle where the Kittiwakes nest (see cover photo). There is no signage and the majority of people using the path will be unaware of the birds nesting metres away. The green right next to the Castle also allows the public to walk virtually right up to the colony.
- The north Harbour entrance can be accessed via footpath in a similar manner to the main Castle, although it is not possible to walk all the way along. The south Harbour entrance is not accessible. Increased marine recreational use of the harbour may also be responsible for the declines in these areas as the harbour entrance is only ~20 m wide.
- The number of nests on the (inaccessible) Leisure Pool cliffs has been gradually increasing. Although numbers of birds nesting in the Inner Castle have also been increasing, over recent years numbers have dropped reducing from ~296 in 2015 to 218 in 2020, although further years data will be required to confirm that it is a genuine decline rather than natural variation.
- Although the magazine is also inaccessible to the public and is located close to the Leisure Pool cliffs, the number of nests in this location has declined over the past decade from around 150 nests in 2010 to only 62 in 2020. The reasons for this decline are unclear. The small number of birds nesting on the rocky outcrops and sea wall has remained relatively constant over the past twenty years, suggesting that these areas may be at capacity.
- In general, there has been a re-distribution of birds away from the more disturbed areas, namely the main Castle and the north and south harbour entrances in favour of less accessible sites such as the Inner Castle (fenced off to the public), the leisure pool cliffs and the rocky outcrops (inaccessible). However, despite this re-distribution, there is still an underlying trend of decline in overall numbers of nests at Dunbar driven by the decline of nests on the main Castle.
3.2.4. Colony productivity
- Productivity data is available for the site from 1990–2015 (Table 3.2, Figure 3.5). Recording stopped in 2015 due to lack of resource. However, the location of the monitoring plot will need to be moved from the main Castle as there are now too few nests in this location (T. Sykes, ELCRS, pers. comm.).
- Average annual productivity over the 25 years is 0.83, which mirrors JNCC productivity data for Scotland from 2009–2019[26]. However, productivity was very low between 2004 and 2008, coincident with the mass appearance of Snake Pipefish Entelurus aequoreus and poor breeding success at a number of colonies nationwide. In these years, chicks starved and mortality rates were 50–60%. In many cases chicks were found emaciated and choking on Snake pipefish, which they were unable to swallow due to their long and cartilaginous structure. In 2013 the colony suffered a very severe storm, which resulted in the loss of many chicks and adults due to exposure (T. Sykes, ELCRS, pers. comm.).
- When fledglings and adults are ringed, data is gathered on their body mass. Between 1993-2007 there was a reduction over time in nestling body mass combined with larger numbers of nestlings dying even though colony size increased. It is speculated that poor prey availability was responsible for declines, although no decline in adult body mass was recorded (Coleman et al. 2011).
- Although Kittiwakes may lay up to 3 eggs, clutch size decreased to just a single egg during the period from 2004-2008. Although the situation has improved, and many birds now lay 2 eggs and go on to successfully fledge two chicks, there are no nests with 3 eggs, again indicating that prey supply is potentially poor (T. Sykes pers. comm.). However, this is not unique to Dunbar, and reflects the situation throughout the local Forth area.
The RED numbers show the number of nests in each area from 2020. GREEN numbers show Kittiwake count areas: | |||
1-main Castle, 2-Inner Castle, 3-South Harbour entrance, | 4-North Harbour entrance, 5-Magazine, 6-Leisure Pool | 7-Johnston’s Hole 1, 8-Johnston’s Hole 2, 9-Rock outcrop 1 | 10-sea wall, 11-Rock outcrop 2, 12-Johnston’s Hole 3 |
Figure 3.2: Location of Kittiwake nesting areas within Dunbar Harbour and surrounding coastline.
Figure 3.3: Number of Kittiwake nests (AON) at Dunbar by location.
Table 3.1: Number and location of Kittiwake nests from Dunbar Castle and surrounding coastline from 1979-2020. Numbers relate to areas shown on Figure 4.2 Data supplied by East Lothian Council[27]
Table 3.2: Productivity monitoring from Dunbar Castle 1990–2015. Data from East Lothian Council.
Figure 3.5: Changes in productivity (number of chicks fledged per pair) from Dunbar 1990–2015.
3.3. Project Description
3.3. Project Description
3.3.1. Stakeholder meeting
- An online meeting was held on 27th December 2021 with the relevant stakeholders (DHT, ELC and ELCRS) to discuss the objectives of the warden, how the warden might interface with the workings of the harbour, working facilities and logistics. The description of the project is derived from this process, and therefore has the in-principle support of all stakeholders. This report outlines very briefly how the wardening would work on a practical level but is primarily focussed on presenting the evidence behind why wardening has been recommended at Dunbar. Further detail on the delivery of the project is located within the Implementation and Monitoring Plan
- It was agreed that the objectives of the warden would be to identify limiting factors and implement solutions to improve both the number of birds nesting at Dunbar and their breeding success. The warden would also work closely with the Dunbar Harbour Trust and engage with both the fishermen and other users of the harbour as well as the general public.
- Given that duties would be beyond what might normally be associated with a wardening post, it was agreed that the term warden/researcher might be more suitable. Although there is information available about the colony, it is not comprehensive and further study will be required to either identify or confirm the actions required. Furthermore, the issues may change over the lifetime of the project so the warden/researcher would need to be able to identify emerging issues, and where necessary gather data and develop solutions.
- The factors believed to be limiting colony success at present are discussed in further detail below. Tackling these factors would, at least initially, constitute the main elements of the warden/researcher’s job.
3.3.2. Human disturbance
- The issue of human disturbance is highly relevant to Dunbar because of the number of people using the Harbour, the range of potentially disturbing activities undertaken, and the decline in the number of Kittiwake nests in areas that are accessible to the general public. Although the impacts of human disturbance have often been dismissed as minor in nature, and therefore inconsequential in relation to the more commonly acknowledged pressures such as prey availability and extreme weather events, there is increasing evidence that suggests human disturbance can cause a measurable and significant level of harm in certain circumstances.
- Although direct mortality of birds due to human disturbance has been recorded in a few circumstances (Yasue & Dearden 2006, Lilley 1999), this is rare. However, there is now a significant body of work that demonstrates reduced breeding success in locations where disturbance is greater (Arroyo & Razin 2006, Ruhlen et al. 2003, Bolduc & Guillemette 2003, Murrison 2002). There are also an increasing number of studies showing how birds view habitats as lower quality if they are regularly used by people (Mallord et al. 2007, Gill et al. 1996, Bötsch et al. 2018, Remacha et al. 2016) and there are many examples of otherwise suitable habitat being unused because of disturbance (Gill 1996, Kaiser et al. 2006, Liley & Sutherland 2007). A few studies have now been able to demonstrate that human disturbance may result in population level impacts (Liley & Sutherland 2007, Mallord et al. 2007, Stillman et al. 2007, West et al. 2002).
- The negative impacts of human disturbance on ground-nesting shorebirds were identified some time ago (Carney & Sydeman 1999) and the benefits of fencing areas to improve the breeding success of ground-nesting shorebirds and terns are widely acknowledged (Verhoeven et al. 2022, Babcock & Booth 2020b). Non-electric fences protect areas from disturbance by humans and dogs (Babcock & Booth 2020b), whilst electric fences also exclude mammalian predators such as foxes (Verhoeven et al. 2022). Wardens are necessary to ensure that people and dogs remain outside of fenced areas, particularly in busy locations, and positive engagement with the public is required to achieve this. People are considered more likely to behave in a reasonable way if they are being observed (Babcock & Booth 2020b). However, the value of fences and wardening for cliff-nesting species are less clear. These species are already nesting in inaccessible areas and there is a perception that little can be done to help them.
- However, there is increasing evidence that human disturbance also impacts on cliff-nesting seabirds. Heart-rate monitors attached to Kittiwakes nesting near the path at St Abbs National Nature Reserve (NNR) showed that birds experienced a rise in heart rate as a consequence of stress when visitors were close, even though no external behavioural response was observable (Beale & Monaghan 2004).
- The results of another disturbance study on Gannet at Great Saltee, an unmanaged island reserve offshore of Ireland, were also surprising. Gannet is generally assumed to be a robust and approachable species and is thought to be tolerant of both humans (presumably because numbers have been increasing at virtually all colonies). Allbrook and Quinn (2020) found that the percentage of successful nests declined with proximity to the edge of the colony. Nestling success averaged 0.33 chicks fledged per nest in the disturbed area compared to 0.52 in the undisturbed control. The study also concluded that visitor proximity to the colony reduced in the presence of an information sign. One of the authors on a single day observed 10 separate incidents of egg predation from gulls caused directly by human behaviour. This demonstrates that human disturbance can also impact indirectly on nest success (Allbrook & Quinn 2020).
- Another recent study has been able to quantify the impacts of human disturbance on colony productivity. A study on European Storm Petrels nesting on Mousa, a small uninhabited island in Shetland, showed that nestling mortality was higher in areas exposed to high visitor pressure and that overall colony productivity was reduced by ≤ 1.6% compared with that expected in the absence of visitors (Watson et al. 2014). The results of this study were surprising because it had always been assumed that Storm Petrels were not impacted by human disturbance. Like many other burrow- and cavity-nesting seabirds, Storm Petrels are only active within the colony at night. During the day, birds either remain in the underground nest, out of sight, or are foraging at sea. Storm petrels prefer dark chambers and nests are usually at least 30 cm below ground and without a direct view to the exterior; therefore, there is no visual contact between human visitors and the storm petrels (Watson et al. 2014).
- These studies demonstrate that human disturbance can impact on seabird productivity, even in remote locations where there is no obvious disturbance problem. By contrast the Dunbar Kittiwakes are subject to various differing and constant disturbance sources.
- Wardens are typically employed to prevent damage to wildlife and to provide information to visitors. They may also carry out other activities such as monitoring, reporting, habitat improvement, repair, dealing with emergencies (fire, flooding, storm damage), litter removal, guided talks, and training and supervising volunteers. They may also be responsible for enforcing wildlife law by passing on details of wildlife crime to the police. The concept of having a warden to look after a seabird colony is not a new one. In 1861 Archdeacon Charles Thorp arranged the purchase of some of the Farne Islands and employment of a warden to protect threatened seabirds. The idea of having a warden to protect animals is derived from the medieval gamekeepers who stopped any intruders from hunting the King’s deer. The concept of having a warden present on site to prevent others instigating damaging behaviour remains unchanged.
- The value of wardening reserves supporting nationally and internationally important numbers of birds is generally acknowledged and relates directly to the conservation value of the species concerned. A need for wardening may also arise in response to conflicts between wildlife and people occupying overlapping spaces. For example, in east Norfolk a local volunteer scheme, the Friends of Horsey Seals, was initiated to reduce disturbance of breeding Grey seals by people and dogs. This voluntary wardening scheme has been extremely successful; seals and people are physically separated by fences and visitors are restricted from the beach and channelled along a path. This has reduced disturbance to the seals, whilst the presence of wardens has created a safer and more informative experience for visitors. The colony has expanded both geographically and in terms of numbers of pups born. Although this increase reflects national trends, the scale of colony expansion at Horsey is still notable.
- The benefits of a warden are clear, and all the activities typically undertaken by a warden would be of significant benefit at Dunbar; the Kittiwake colony has a problem with fishing litter, engagement with the local community is required to foster a better sense of ownership of the colony in general, the site would benefit from habitat enhancement, and a warden and volunteers would be able to gather information on the birds, which is currently lacking. The presence of a warden would prevent some of the behaviour that at times results in police involvement. With a warden visitors can be kept away from the immediate vicinity of the colony, but in return receive a more informative experience by talking to a warden, learning about the Kittiwakes and even seeing nests and chicks through a telescope. The Kittiwake colony is so close to the town and working harbour, which is also a busy tourist spot, that a permanent warden is considered a necessity in safeguarding the colony and ensuring the site’s success in future years.
- The East Lothian Visitor Survey carried out in 2021 showed that Dunbar is second most visited town in East Lothian (after North Berwick) and was visited by 40% of tourists staying in the area (STR 2022). Dunbar Harbour is a scenic and historic site supporting not only the Castle, but an old fort (The Battery), a converted granary (now the Dunbar Harbour Trust), and a magazine where ammunition was historically stored. Dunbar has previously been the recipient of a £4 million grant from the Heritage Lottery Fund to support the regeneration of the historic urban environment (https://www.shbt.org.uk/our-projects/dunbar-townscape-heritage-initiative/).
- The DHT was incorporated in 1999 with the objective of ‘operating the harbour for the benefit of port users and the local community…as well as becoming a focal point for the local community’. The DHT has implemented various projects to improve the area and improve access to the Harbour and to enhance appreciation of its historical features. The largest of these projects was renovation of The Battery in 2017. This project has transformed this ruined fort (located ~200 m from the Kittiwake colony) into an outdoor events venue that now hosts regular music and theatre events as well as community festivals. There is now a Dunbar Battery Theatre Company, and an aspiration to increase its use and make Dunbar Harbour a ‘go-to’ place in East Lothian for the performing arts. The impacts of these events on the Kittiwakes have not been monitored, although it is speculated that the renovation of The Battery may be associated with the suspected decline in nesting Kittiwakes in the Inner Castle in recent years.
- Offshore recreational use of the Harbour and surrounding waters for water sports is also increasing and activities undertaken locally include kayaking, paddle boarding, diving, sailing, coastal rowing and surfing. The impacts of these various activities on the Kittiwakes are also unstudied.
- Use of the harbour is thought to have increased significantly as an area for walking both by locals and visitors, especially over the last two years It is understood that visitor numbers have trebled due to the popularity of ‘staycations’ during the SARS-Cov-2 pandemic when overseas travel has been restricted (T. Sykes East Lothian Countryside Ranger Service pers. comm.). This reflects general trends observed in both East Lothian and the UK during the pandemic for increased uptake of outdoor activities such as hiking, outdoor swimming and trips to the beach in preference to indoor attractions (STR 2022).
- On occasion, local children throw stones at the Kittiwakes nesting on the main Castle. This may be especially damaging if the disturbance takes place early in the season before eggs are laid. If the ELCRS is aware and the children responsible are known locally, the police are informed and will talk to parents. If children are not known, it can take longer to resolve. The simple presence of a dedicated site warden is likely to stop such behaviour and would also stop local youth from ‘tombstoning’ (jumping from height into the sea) from near the Kittiwake colony, another activity that is contributing to disturbance in the area.
- The importance of educating the local children in school is critical in stopping anti-wildlife behaviour and would go a long way in engendering a sense of connection to and ownership of the Kittiwake colony. A warden would be able to undertake visits to local schools to teach children about the lifecycle of the Kittiwake, perhaps coupled with a follow-up trip to the colony to show children nesting birds and chicks using a telescope.
- During the main tourist season a warden could be on site, perhaps at the Battery, at specific times with a telescope to provide opportunities for both visitors and members of the local community to see the birds in a way that is not normally possible and to learn more about them.
- Increased vessel movements and people in the harbour can reduce Kittiwake foraging activity. It is known that the Kittiwakes favour feeding and loafing at Belhaven Bay at low water and may be displaced by people and surfers if the beach is busy. In the past, council led social media campaigns have been successful in making the public aware of these types of issues, and it is suggested that this could be taken up again and continued by the warden. Codes of conduct could be developed with local groups to reduce the potential impacts of these activities.
- Changes in the distribution of nests and the decline in nests on the main Castle and harbour entrances strongly suggests that human disturbance may be causing direct displacement of birds away from key nesting areas, and without intervention it is likely that the current trend of decline will continue. Once colony size decreases beyond a certain level then the birds are more vulnerable to predators, and colony success decreases.
- The path that runs along the main Castle colony and the green in front of the steps are the key areas where there is greatest overlap between the public and the Kittiwakes. At present people, dogs and children can walk along the narrow path that directly abuts the outer wall of the main Castle where the Kittiwakes nest (see figure 4.6). There is no signage and people walking along this path may very easily and completely inadvertently disturb the birds unaware of their nests just meters away. The green right next to the Castle also allows the public to walk right up to the colony.
- How birds respond to disturbance is context and species specific and birds may habituate to certain activities over time. However, judging habituation is problematic; for example, in the St Abbs Kittiwake study described above (Beale & Monaghan 2004) the birds did not exhibit any outward response to human disturbance, and it could easily be assumed that they had successfully habituated to the presence of humans on the path. How disturbance could be monitored and assessed at Dunbar is discussed further in section 4.3.7.
- Further evaluation is needed to determine how the various types of disturbance may be impacting on the birds, and to establish how these may be tackled. Disturbance could be reduced by restricting access to the front face and entrance stepway on the south side of the harbour. Fencing off the green in front of the steps to add a buffer between the Kittiwakes and the public would also be of significant benefit (T. Sykes, pers. comm).
- Although some of this information is anecdotal by nature, there is reasonable evidence that disturbance is a significant factor at this colony. Lack of targeted data is inevitable as there is currently no funding mechanism to enable information to be collected officially. However, the staff who have supplied information to support this project are experienced professionals and the prospect of having a Kittiwake warden is well-supported locally amongst the relevant stakeholders.
3.3.3. Prey
- Although the studies cited above show that human disturbance can potentially impact on colony productivity, it is important to evaluate the role of other known influential factors. JNCC Seabird Monitoring Programme data indicates that Kittiwake numbers increased by around 24% between the late 1960s and the mid-1980s possibly due to the cessation of egg collecting and hunting (Cramp et al. 1974). However, catastrophic declines followed from at least 1986 onwards, and there are now around 50% fewer birds than in the late 1960s (JNCC 2021). Changes in the marine environment due to warmer sea temperatures and the associated decline in the abundance of sandeels is thought to be responsible (Frederiksen et al. 2004). On this basis, poor prey availability due to climate change is routinely identified as being the main pressure affecting this species.
- Unlike the other Scottish colonies, the number of Kittiwakes nesting at Dunbar increased during the 1990s (Figure 3.1). Productivity data is available for the site from 1990-2015 and over the 25-year period averages 0.83, mirroring the JNCC productivity data for Scotland from 2009-2019. Although productivity is not a direct proxy for prey abundance, there is no apparent evidence to suggest that foraging conditions differed significantly at Dunbar to any of the other local colonies. As mentioned previously it is likely that the increases observed at Dunbar during the 1990s relate to the closure of the Castle to the public resulting in increasing numbers of Kittiwakes occupying the main and Inner Castle areas.
- Recent analysis assessing the benefits of the existing sandeel fishery closure (implemented in 2000) on local Kittiwake colonies shows that even though the Isle of May and Dunbar are only 40 km apart, breeding success for the Isle of May has increased by 17% since the sandeel fishery closure within the Firth of Forth as offshore foraging conditions have improved, whilst the trend of moderate decline has continued at Dunbar. It is hypothesized that human disturbance is the cause for the differing trends observed at these two colonies (Searle et al. 2022 in prep.). If prey was the limiting factor, then similar increases would be expected at Dunbar to those observed on the Isle of May. The decline at Dunbar suggests there are other local factors affecting the success of this colony, the most obvious of which is human disturbance (see Section 3.3.2).
- As a general principle, the potential impacts of poor prey availability on colony success must be acknowledged. High Sea Surface Temperature (SST) and poor prey availability, as experienced during 2007 and 2008, resulted in extremely low productivity levels across many Kittiwake colonies. The re-occurrence of similar conditions could negatively impact on the delivery of conservation targets at Dunbar. However, the effectiveness of colony-based measures at any site could be affected by these unpredictable large scale climatic events.
3.3.4. Predation
- Since the prey situation has gradually ameliorated within the Firth of Forth over the last decade, the main cause of chick mortality for Kittiwakes appears to no longer be starvation, but predation by Herring Gull with mortality estimated at ~5% (T. Sykes, ELCRS, pers.comm.). Assuming a colony size of 808 nests, an average of 0.83 chicks fledged per nest, tackling Herring Gull predation would save ~34 chicks per year. However, the indications from stakeholder engagement are that any work that may impacts negatively on Herring Gull in any way will not be viewed favourably.
- Rats are also thought to have become a problem in the main Castle. This is believed to be because fishermen have been stacking up old creels next to the castle wall (Figure 3.6). The creels attract rats, which are then able to climb up the creels and predate Kittiwake nests in this area, which is also where the productivity monitoring is carried out. As a consequence, Kittiwake numbers in this area are declining, signifying that the value of the habitat has decreased. The favoured nesting area is now the Inner Castle. The lack of storage space for the fishermen at Dunbar Harbour is an ongoing issue and removing the creels and implementing a rodent control programme over the winter months would be effective in reducing rat numbers and improving the value of the habitat for Kittiwakes.
3.3.5. Habitat enhancement
- It was suggested during the stakeholder consultation meeting with DHT,ELC and ELCRS that the site could benefit from habitat enhancement. The need for habitat enhancement was questioned by NatureScot on the basis that there is spare nest capacity with possibly 300-500 available[28].
- This availability of physical nest space is undisputed, and the estimate of available space seems sensible on the basis that the colony has declined by 400 nests over the past 20 years. In view of the disturbance issues, it is intuitive to focus on reducing disturbance around the main Castle area and harbour entrance, and in so doing encourage the birds to return to these nesting areas.
- The ruined castle naturally provides ledges, overhangs and shelter, and parts of it (the main Castle and Inner Castle particularly) are considered optimum habitat. The Kittiwakes nest in the cavities left by lost masonry, which has resulted in a network of sheltered nesting areas, many of which benefit from overhangs and which offer more shelter than many natural cliff faces (Figure 3.7).
- However, nest numbers are declining on the main Castle front and birds are instead nesting in some locations that are considered sub-optimal, particularly the sea wall where nests are always flooded out.
- However, it is possible that recovery may take time and given that the site suffers from significant levels of disturbance from multiple sources, it may not be possible to tackle all disturbance sources effectively. It is possible that adding extra nesting ledges to areas which are not disturbed but which may already be at full capacity, such as the rocky outcrops, could be an effective way of helping Kittiwake numbers to recover.
- It has also been suggested that changes in prevailing wind direction in favour of more north-easterly winds may affect choice of nest site, and if this is the case then other nesting areas may now be more favourable. Although further study would be needed to confirm whether this is the case, habitat enhancement could provide scope for adding nesting ledges in areas that are currently preferred.
- A better understanding of productivity across the different sub-colonies is needed to understand how successful each of the sub-sites are before habitat enhancement can be considered. To date productivity has only been monitored on the main Castle. It is understood that the small numbers of birds nesting on the outer sea wall routinely fail as the nests are too low and they are washed out in storms and/or high tides (T. Sykes pers. comm). Productivity data from other locations such as the rocky outcrops, the magazine and the cliffs are needed to assess whether similar problems are occurring and whether habitat enhancement in the form of more ledges higher up would be of benefit in these specific areas.
- In general, it is felt that there is scope to carry out habitat enhancement in the future at Dunbar in a manner that will benefit the birds through the provision of additional nesting habitat in more favourable locations. However, a better understanding of the site is required to reduce uncertainty and to ensure that any work planned has the best chance of success.
- The Magazine has the potential to support more nesting birds if the colony increases. It is also considered to be the area that would most benefit from habitat enhancement. Other areas that would also benefit include Rock outcrop 1 (9), Johnston’s Hole 1 (7), Rock outcrop 2 (11), North Harbour Entrance (4), Johnston’s Hole 3 (12), and Johnston’s Hole 2 (8)[29]. These areas, shown on Figure 4.8, do not contain the same types of tall walls, ledges and overhangs offered by the main castle.
- Habitat enhancement would involve adding ledges and overhangs in these areas. The overhangs need to be sized correctly to prevent Herring Gulls from landing and predating nests. Nests that occur naturally with overhangs are routinely observed to be successful on the Isle of May for this reason (F. Daunt, pers. comm).
- In view of the historic value of the site, further liaison will be undertaken involving the local Conservation Officer to agree an acceptable plan for the work. These areas are also challenging to access, so skilled contractors will be employed by the Applicant.
3.3.6. Fishing litter
- Dunbar appears to be one of the worst Kittiwake colonies for plastic debris (Figure 3.9). A study looking at the presence of plastics in seabird nests examined 3,681 Kittiwake nests from 33 colonies. Only 4% of nests contained plastics, with most colonies containing no debris. The two exceptional colonies in the study were Dunbar and an oil rig in the North Sea, with 31% and 49% of all the nests respectively, containing anthropogenic debris (O’Hanlon et al. 2021). This reinforces comments made during stakeholder consultation that nearly all of the nests at Dunbar have plastic incorporated within them, and that fishermen regularly discard offcuts of net directly into the Harbour.
- Although it is understood that the DHT and the Harbourmaster have been working to stop this practice and have also introduced the Fishing for Litter initiative[30], it would be helpful to continue and build on this work to ensure that higher standards are attained and maintained.
- Since Kittiwakes re-use nests year on year, and the plastic is integrated within the nest (Figure 3.9), totally removing all plastic would result in the nests falling apart. Therefore, it is suggested that the danger of entanglement could be minimized by clipping any trailing net or rope. Any available small pieces of plastic that could be ingested would also be removed. It is known that a small number of birds (often adults) perish every year due to entanglement, and this could be avoided by removing the anthropogenic debris. It is harder to know how many birds ingest plastic, but those that do are likely to suffer negative impacts in the longer term (see Section 7.4). The warden would be able to remove debris from most of the nests, although a climbing crew would be required for some of the difficult areas. A warden would also be helpful in ensuring that the colony remains debris-free into the future.
- It was agreed during stakeholder meetings that a representative of the fishing community should be involved in further discussions regarding the Kittiwake warden so that they are kept aware of what is happening within the Harbour, have a clearer understanding of what the issues are and also what will be expected from them and why.
3.3.7. Wardening role
- This section broadly describes the principal components of the wardening role. Full details of how the work would be implemented are presented within the Implementation and Monitoring Plan.
- Since human disturbance is a key pressure, gathering more information on the different types of human disturbance and how the birds respond is considered a priority. Table 3.3 shows the type of approach that could be undertaken to tackle key questions.
- Community engagement is central to the success of this work, and an approach that seeks to understand why people are visiting Dunbar Harbour and that takes their needs into account (alongside those of the Kittiwakes) is considered more likely to result in long-term success. This approach would also accommodate the goals of DHT, who would be an important stakeholder in this project.
- The need to monitor and quantify the success of work undertaken under this project is understood and is in contrast with other wardening work, which is generally undertaken solely for the purposes of conservation. The need to gather scientific data on the warden and how their presence may (or may not) reduce disturbance in order to establish the benefits to the birds is a novel idea, although other studies provide a precedent (Liley & Panter 2017).

- In addition to human disturbance, it is anticipated that the warden would also gather colony counts, map nesting locations and monitor productivity in a number of different locations. Cameras and monitoring equipment could be used to gather data on seabird demographics (attendance rates, chick provisioning rates etc). It is anticipated that the warden would also identify and find mechanisms of tackling other pressures that may be impacting on the birds, such controlling rodent numbers, fishing litter and other items mentioned above. Any major changes or decisions relating to the project will need to be evidence based as it is acknowledged that the needs of the site may change throughout the project lifespan.
- Since one of the fundamental aims of the project is to foster a better sense of ownership of the colony amongst the local community, undertaking educational visits to schools and public engagement will also form a key element of the work for the warden.
3.4. Conservation targets
3.4. Conservation targets
- At its peak in 2000 the colony numbered just short of 1,200 pairs. It is speculated that bringing the colony back to this level from the 808 nests observed in 2020 would be a realistic conservation target of ~400 pairs (800 birds). This works out at 23 birds per year assuming a 35-year project lifespan. Although recovery is unlikely to occur in this linear fashion as seabird counts always fluctuate, the conservation benefit is framed in these terms to allow comparison with potential mortality estimates for the Proposed Development.
- This conservation target seems feasible on the basis that the colony previously supported >1,100 pairs during several years during the noughties (2006, 2007 and 2010). It is thought likely that the birds will recolonise the Castle if it is fenced off to minimise human disturbance, the creels are removed, and the rodents brought under control. If birds are able to nest in the best habitat, then productivity would also be expected to improve.
- After the Castle and Inner Castle, the Magazine is considered the next best nesting area. If habitat improvements are carried out at the Magazine, then it has the potential to support many more pairs, which would theoretically enable the colony to expand beyond 1,200 AON. Improving the Magazine and the other sub-optimal areas that are currently in use will increase the productivity of any pairs that continue to nest in these locations.
- Although the nests are generally well consolidated, some trailing net and plastic results in mortality though entanglement. Entanglement is believed to be a cause of death for a few individuals per year (both adults and chicks are susceptible). The risk posed by ingestion of plastic is harder to quantify (see Section 7.4 for further discussion), but a cleaner harbour will reduce the quantity of plastic ingested by foraging birds in the waters around the colony.
- It was suggested by MSS following the Compensation Consultation meeting on 8th June 2022 that the compensation benefits resulting from the work at Dunbar should be set within the context of predicted mortality impacts from both the Proposed Development and other wind farm sites within the Forth[31]. Although this is a valid consideration, it is not possible as Dunbar is included in a ‘non-SPA’ total count within the assessment for the Proposed Development, and numbers are not presented for it as a colony in any of the other EIAs of local offshore wind farm sites (e.g. Inchcape, Seagreen ,or Neart Na Gaoithe).
3.5. Timescale & mechanism for delivery
3.5. Timescale & mechanism for delivery
- The mechanism for delivery is presented in full in the Implementation and Monitoring Plan and only an overview is provided here.
- There is local support for the work with positive engagement from both ELC and DHT, and there are no reasons why implementation of the measure cannot be rapid.
- It is suggested that the Dunbar Kittiwake warden would employed through the ELCRS (part of East Lothian Council) with the post funded by the Applicant, although they would also work closely with the DHT and other local stakeholders.
- The items related to the general protection of the site could start from mid-March when the birds return, as could the education initiatives, improving public awareness of the colony through social media, and liaison with the fishermen regarding fishing litter. Much of the data gathering, for example counts, productivity, provisioning rates, and predator monitoring could also be carried out on the birds return in mid-March.
- Rodent control should be done during the winter and could be implemented over the winter when the birds are not present. Similarly, removal of plastic from nest sites needs to be carried out over the winter, and liaison with the Council and Conservation Officer will be required beforehand due to the building’s historic status. It would also be necessary to employ contractors to access the more difficult nesting areas.
3.6. Additionality & uncertainty
3.6. Additionality & uncertainty
- Since Dunbar is not designated for nature conservation, none of the recommended improvements would happen without an additional external funding source. Although the council have maintained a responsibility for counting the Kittiwakes, there is no funding to deal with any of the other aspects described, and productivity monitoring was stopped in 2015 due to lack of resources. Therefore the only item that is already being carried out at the time of writing are the annual bird counts.
- The DHT states that one of their objectives is to improve the environment within the harbour. It could be argued that this does overlap in a minor way with the role of the Kittiwake warden, particularly with regards to liaison over fishing litter. However, the DHT is a charity and its resources are also limited. A dedicated warden would be helpful in reinforcing these messages and would be able to ensure that better standards are attained and maintained in the longer term.
- Whilst a conservation target of 800 birds seems reasonable, it is uncertain how long it will take to achieve given that the factors limiting the success of the colony are not completely understood. However, it appears that the issues are colony-based rather than prey-based (Searle et al. 2022 in prep) then it is possible that the response could be rapid if disturbance can be tackled successfully.
- Although the measures are tried, tested and deliverable as they represent standard conservation management practice, the need to monitor their success in order to quantify the benefits of the project is not standard practice, although other studies set a precedent (Liley & Panter 2017). Dunbar Castle is a Scheduled Monument, and Dunbar Harbour is a historic conservation area. With careful design and the use of non-invasive installation techniques, the Applicant does not anticipate the need for a licence. If a licence is required, an application would be made to HES. However, as non-invasive installation techniques would be used to avoid any structural damage to the scheduled monument and these techniques would be agreed with HES, as well as the fact these measures has support from the landowner, the Applicant does not foresee any impediments to gaining such a licence if it was required.
- There is some uncertainty regarding the scale of human disturbance impacts at the site, and also to what extent disturbance can be minimised. There are no obvious issues with fencing off the green so that there is a buffer area between the public and the breeding Kittiwakes. In addition, DHT have confirmed that the fishermen do not have the right to store creels at the front of the Castle, although this does require their active engagement and willingness to reduce disturbance to the extent that is desired.
- It is possible that the Kittiwakes may be cautious about returning to the Castle, even if disturbance is reduced, as birds typically return to the same place to breed. However, it is anticipated that young birds nesting for the first time would fill in the area should this occur.
- Educating children and the wider public may not directly lead to an improvement in the success of the colony. However, it is still considered of essential importance if Kittiwakes and humans are going to co-exist successfully in such close proximity into the future.
- Similarly, it may be difficult to quantify the benefit of removing trailing net/rope and small potentially ingestible plastic items from the nesting area (and general harbour). However, the removal of these plastics follows the holistic principles set out in the main Derogation case whereby compensation is potentially strengthened by tackling multiple limiting factors whenever possible.
- In summary, the provision of a site warden and potential management of disturbance and any other factors that may be negatively impacting on the Dunbar Kittiwakes is additional to the work currently undertaken at this site, which is limited to annual colony counts. Since recent research indicates that the issues at Dunbar to relate to colony management rather than prey (Searle et al. 2022, in prep.), then there is reasonable certainty that the project would be effective in improving the success of this locally important colony.
4. TIER I: REDUCTION OF THE GANNET HARVEST AT SULA SGEIR
4. TIER I: REDUCTION OF THE GANNET HARVEST AT SULA SGEIR
4.1. Background
4.1. Background
- The Gannet population in the UK has risen by 34% between surveys carried out in 1969/70 and in 2013–15 (JNCC 2021). This increase is attributed to the cessation of hunting[32] dating from the end of the 19th century (Nelson 2002). However, Gannet is classified as being of ‘Amber’ conservation concern because the UK contains an internationally important breeding population (at least 20% of the European population) and has at least 50% of breeding birds present in 10 or fewer colonies (Eaton et al. 2015).
- Bass Rock supports the largest UK colony and is also the largest gannetry in the world supporting 14.3% of the world population (Murray et al. 2014). The population numbered just 8,077 pairs in 1968 and increased by 832% to reach 75,259 pairs in 2014. The rate of increase at 4.4.% per annum (from 2003/4) has been higher at Bass Rock than is typical at around 2% per year (JNCC 2021).
- All Gannet SPA breeding populations are in Favourable conservation status which is in marked contrast to the situation for many other seabird populations (Murray et al. 2014). The Outer Firth of Forth and St Andrews Bay Complex SPA was recently classified (December 2020) with breeding Gannet as a breeding season feature (as was The Seas off St Kilda SPA), which effectively recognises the area as of importance for foraging Gannet, mainly originating from Bass Rock. No offshore sites were listed as being designated for non-breeding Gannet in the 3rd UKSPA review (Stroud et al. 2016).
- Gannet productivity has been stable over time (JNCC 2021) suggesting that the population is generally not limited by prey shortages due to their adaptability and ability to forage over great distances (typically up to 700 km) from their nest (Hamer et al. 2000). This conclusion also logically accords to steadily increasing populations in the UK. However, extended foraging range is also a response to reduced prey availability perhaps as a result of intense competition for resource around large colonies in particular. Thus, Gannets from Bass Rock travel as far as the Bergen/Viking Bank near the Norwegian coast to find food, a round trip of up to 1,291 km (Hamer et al. 2007). Usually, one parent will stay with a chick whilst the other forages, though if left for long enough it will eventually leave to find food. This leaves the chick vulnerable to attack, mainly from other Gannets seeking prime nesting space. Impacts upon breeding success are anticipated if reduced prey availability promote any further increases in trip duration or foraging effort.
- Fisheries-based measures to reduce or remove sandeel fishing pressure in SA4 will be of benefit to Gannet, as they forage on sandeels as well as larger fish such as Atlantic Mackerel Scomber scombrus, gadoids and clupeids. A study of 266 regurgitate samples from Gannets at Bass Rock showed that the main prey items in terms of frequency of occurrence were Mackerel, sandeels (mainly 0-group), Sprat and Herring. In terms of biomass, sandeel accounted for 17.9% of diet (Hamer et al. 2000). Fisheries Compensatory Measures (FCM) are discussed in a separate FCM Report submitted alongside this document.
- The possibility of compensating for Gannet within the area in proximity to the Proposed Development was assessed in ECON & ABPmer (2021a). Although Gannets are not limited by prey, they are limited by habitat availability at Bass Rock as the colony is virtually at full capacity. Under these conditions, it is likely that many chicks (perhaps even several thousand per year) will be lost falling from cliffs due to competition for space with other Gannets. A 3-year study at Ailsa Craig, a colony where, unusually, any falling gannets can be retrieved from scree underneath the cliffs, recorded between 445–461 dead or injured gannet chicks over the course of a breeding season, accounting for about 6% of the chicks hatched (Wanless 1983). The author postulates that this number may be reduced at sites where birds fall into the sea, as some fledged birds may survive the fall. However, even taking this into account the losses are likely to be substantial. The possibility of establishing another gannetry was discussed with stakeholders but was felt to be problematic due to lack of other suitable nesting cliffs within the Forth coupled with the natural expansion of birds into St Abbs NNR, where a few pairs have started nesting on one of the stacks in recent years.
- Reducing the impacts of predation and human disturbance on Gannet were also considered. Whilst a few Great Black-backed Gulls are known to take Gannet eggs at Bass Rock, the size of the gannetry is such that this is not a significant problem. However, at other sites such Sule Skerry, which in 2003 was a start-up colony of 15 nests, 30% of nests were predated by Great Black-backed Gull. However, even under these relatively difficult conditions this colony continued to increase year on year and by 2015 held 1,870 nests (Mavor et al. 2004). Since most gannetries are in remote locations, often on inaccessible cliff faces, Gannets were in general felt to be less impacted by human disturbance than other species. Although individual nests at Bass Rock near the path may be disturbed by visitors, the benefits of limiting disturbance are considered likely to be minor.
- Measures to reduce bycatch of Gannet were also evaluated. In summary, Gannet has been identified as one of the seabird species most at risk of bycatch during the breeding season and in the inshore waters of Scotland (Bradbury et al. 2017), with most of the deaths attributable to offshore static net fisheries and longline fisheries (Northridge et al. 2020). In the Forth and Tay area specifically, there is no evidence of significant long-line and gillnet fishing effort that may be impacting local breeding birds. Where longlining does take place it is mainly a more artisanal fishery using feathers rather than baited hooks. Furthermore, the UK Plan of Action on Seabird Bycatch is already collaborating with multiple stakeholders to identify priority measures to reduce or eliminate incidental bycatch. This may translate into feasible pathways that could constitute compensation for wind farm development in the future.
- There is also considerable albeit largely unquantified bycatch of Gannet when overwintering along the Atlantic Iberian coast including Portugal (Oliveira et al. 2015, Calado et al. 2020) and into West Africa (Grémillet et al. 2015, 2020). Gannet is taken especially by longlines, including in artisanal fisheries but also in purse-seines, and generally appears to the most frequent seabird captured. Bycatch has led to a precipitous decline in the number of tagged birds returning to a colony in Brittany, northern France (Grémillet et al. 2020). Losses to fisheries will also invariably affect UK SPA populations although the scale of the impact is currently unknown. As a result, the benefit of implementing measures to reduce bycatch of wintering birds to benefit UK SPAs cannot currently be quantified. Implementation would also require significant international cooperation and seems likely to be extremely difficult.
- By contrast, human predation, in the form of the traditional licenced summer harvest of Gannet at Sula Sgeir results in up to 2,000 fully-grown chicks (known as guga) taken annually. Reported numbers are also close to this limit, with an average of 1,917 taken per year from 2004 to 2014 (Trinder 2016). The disturbance to the whole colony during this process is also significant. Although simple population modelling indicates that the harvest is sustainable, the harvest has reduced the rate of population growth at Sula Sgeir relative to other colonies (Trinder 2016). It also may be the case that harvest affects the growth rate of other Gannet colonies in the region due to natal emigration between colonies. The Sula Sgeir population may therefore act as a sink for emigrant Gannets from other colonies and ending the hunt could lead to increased growth of the surrounding colonies.
- For this reason, reduction of the annual Gannet hunt has been identified as being the most feasible mechanism to compensate for potential losses of Gannet. Due to the highly mobile nature of Gannets, it is anticipated that there would be connectivity between the Bass Rock population (located in proximity to the Proposed Development) and the Sula Sgier colony. Even though, like most seabirds, the majority of Gannets are thought to return to their natal colony to breed, there is evidence that some birds move between colonies as shown by a chick from Eldey (Iceland) that was subsequently found breeding on Bass Rock (Wernham et al. 2002).
4.2. The Annual Gannet Harvest
4.2. The Annual Gannet Harvest
- Historically, the eggs and chicks of a number of seabird species would have been eaten for subsistence by local communities within Scotland, Ireland and the north of England. The annual harvest of Gannet chicks from Sula Sgeir (part of the North Rona and Sula Sgeir SPA) is carried out by a few individuals of the Isle of Lewis some ~65 km to the south and is a traditional practice of significance to cultural heritage mirroring similar seabird harvests elsewhere, such as the Gannet harvest undertaken annually at Mykines in the Faroe Islands. Seabird harvests also continue in other northern nations such as Iceland, Greenland and Norway.
- The annual guga harvest is the only remaining licenced seabird harvest in the UK. Other subsistence hunting of seabirds came to an end in 1954 when the Protection of Birds Act was passed, which banned the killing of seabirds across the whole of the UK. An exception was made to enable 10 men from the village of Ness (on the Isle of Lewis) to apply for permits to cull 2,000 guga annually from Sula Sgeir.
- The harvest involves 10 men from Ness undertaking a long and treacherous journey to Sula Sgeir, which was traditionally carried out on a rowing boat, but now is undertaken on a chartered fishing boat. The Gannet chicks are harvested during the 2-week window before fledging, as they are only considered to be good to eat if taken during this period. On return to Lewis, guga can only be purchased directly from the crew.
- The harvest itself is believed to date from the 16th century, and the men carrying out the harvest today still use more or less the same methods and keep the same traditions. Also, each man has a cairn and places a stone there for each year that they undertake the harvest. On their last visit they place a stone cap on the cairn (Day 2010).
- Historically, Gannet would have been an everyday food item for the men of Ness, particularly over the winter period when fishing was dangerous. Although the passage to Sula Sgeir is treacherous and climbing down the cliffs would have been highly dangerous, fewer men died during the guga hunt than they did fishing. The guga harvest was also considered more reliable than crofting, as crops could fail or suffer blight leaving the community short of food (Murray 2015).
- It used to be that only native men from Ness who could speak Gaelic were allowed to go on the guga hunt, but rules have relaxed since there are so few young people that speak Gaelic. In fact, the population of Ness has halved in the past 50 years, with many young people opting to leave the island due to the lack of opportunity and poor pay associated with crofting for a living.
- The licence has been gradually reduced from around 3,500 guga to 2,500 and is now set at 2,000. Return rates (supplied by NatureScot) indicate that numbers of chicks taken over the past decade ranges between 1723-2000 (Table 4.1).
Table 4.1: Numbers of guga harvested 2011-2021.
Source: Data provided by NatureScot
- This represents some 17.8% of the single chicks from the 11,230 AON recorded in 2013. It is likely that Sula Sgier acts as a population sink for emigrant Gannets from other UK colonies, which are on average growing at around 2% per annum. With the harvest, the rate of population growth at Sula Sgeir appears to be depressed relative to that recorded at other colonies (Figure 4.1).
4.3. Benefits of reducing the Harvest
4.3. Benefits of reducing the Harvest
- Stopping the Gannet harvest is increasingly being viewed as a means of compensation for potential wind farm losses at UK Gannet SPA colonies (MacArthur Green 2021). Wanless et al. (2004) shows that the rate of increase in colony size in relation to breeding numbers in 1969 demonstrates that the rate of growth at Sula Sgeir is far below the expected rate, whereas by contrast Bass Rock lies well above the expected rate (Figure 4.1). Wanless et al. (2004) also point out that in 1969 Sula Sgeir and Bass Rock were of similar size (differing only by 13 AON), but in 2004 Bass Rock contained 5 times as many birds.
- This data supports the hypothesis that Gannet numbers on Sula Sgeir have increased less than at other colonies, indicating that this harvest has apparently affected the rate of colony growth. The harvest of 2,000 chicks per year also probably affects productivity of unharvested nests through the human disturbance involved. Significantly reducing the harvest is considered highly likely to result in the growth of the colony and other colonies in the northeast Atlantic metapopulation of Gannet.
- Halving the quota to 1000 gannets would result in the addition of 258 adult birds to the population (based on the mean survival rates presented in Horswill & Robinson 2015). The impact of adding these birds to the population would need to be modelled to fully demonstrate the benefit that halving the quota would have. However, this calculation demonstrates that in principle this scale of reduction would compensate for the modelled impacts of the Proposed Development.
4.4. Timescale & mechanism for delivery
4.4. Timescale & mechanism for delivery
- The principle for implementing this measure would be based around offering a compensation payment to the guga hunters in return for them reducing their quota. This would not necessarily require any sort of change to the licence and could therefore conceptually be implemented relatively rapidly.
- If a change to the licence is required, then this could take longer. However, the licences are issued annually and therefore any delays would still only be of limited duration.
4.5. Additionality & uncertainty
4.5. Additionality & uncertainty
- The guga hunt is opposed by a number of animal rights groups that have been campaigning for it to cease. For example, the Scottish Society for the Prevention of Cruelty to Animals (SSPCA) claims that the hunt breaches animal welfare laws.
- Opposition to the hunt intensified following Ness Football Club organising a ‘World Guga Eating Championship’ in 2013. One online petition to the hunt reached 70,000 signatures, and in 2017 several of the hunters received death threats.
- Whilst there are many that would like to see the hunt stopped, NatureScot (at the time SNH) released a statement saying that they were satisfied that the method used to dispatch the birds is humane and the hunt is sustainable.
- At the time of writing, there is no evidence of any action currently underway to stop the hunt, although it has been flagged as a means of compensating for potential losses of Gannets from offshore wind farm developments (MacArthur Green 2021). However, there are no known plans to stop the guga hunt for conservation (or any other) purposes.
- Since the harvest is controlled by humans, then there is high degree of certainty that were the current quota were to be halved then the correct number of adult birds would be ‘saved’ to compensate for the potential impacts of Berwick Bank.
5. TIER II: FORTH ISLANDS INCURSION HUB
5. TIER II: FORTH ISLANDS INCURSION HUB
- Section 2.1 describes the background to the EU Biosecurity for LIFE Project. One of the aims of this project is to make biosecurity part of ‘business as usual’ at island SPAs. Previously biosecurity has not been part of routine colony management, although the intention is that by the end of the project biosecurity procedures will be incorporated and carried out routinely (T. Churchyard, RSPB, pers. comm.). However, at present there is no mechanism to fund dealing with re-incursions of invasive species once the EU project comes to an end in July 2023, although the RSPB are in the process of developing incursion response hubs to deal with incursions whilst the project is in operation.
- It is hoped that following the end of the EU Biosecurity for LIFE Project that biosecurity procedures will be incorporated into colony management plans (T. Churchyard, pers. comm.). However, once the EU project ends, there is no mechanism to fund the implementation of additional work at SPAs, such as surveillance monitoring to detect potential incursions or funding to dealing with any incursions of invasive species that may occur in future. This problem applies to all SPA sites, the Forth Islands SPA being the closest to Berwick Bank offshore wind farm.
- Whilst checking wax blocks for signs of rodent chew marks may be relatively easy for a warden to combine with other routine site visits to carryout bird counts, it is harder for SPA managers to fund the eradication of rats in the event of an incursion as the costs may be significant. This lack of funding is of significant concern as maintaining biosecurity is an essential component of ensuring that the Forth Islands SPA remains in favourable condition in the long term. Re-invasion is a serious issue for several of the key seabird islands, namely Craigleith, Lamb and Fidra, all of which are 1.5 km or less from the mainland and as such are in easy range for Brown Rat, which can swim distances of up to 2 km.
- Incursions can and do happen regularly. Since the initial attempt at eradication, rats have already re-colonised Lamb and further eradication effort is again required over the coming winter (T. Churchyard, RSPB, pers. comm.). This island is particularly challenging to monitor regularly as access is difficult, and work is frequently undertaken via a collaboration with a local kayak club. The possibility of the Applicant funding an incursion hub was therefore raised for discussion with stakeholders during the Applicant’s Compensation Consultation process.
- It was suggested that the Applicant could supply funding for an incursion response hub which could be used by any of the Forth Islands (including Inchcolm) to deal with incursions of either rats or any other invasive mammalian predators (e.g. stoat, mink etc). An incursion hub would ensure that these islands would remain rodent free in the long term. The impacts of rats on nesting seabirds are described in detail in Section 2 and on this basis keeping seabird islands free from rats is considered an important component of maintaining favourable condition status for the Forth Islands SPA.
- Conceptually the project would involve sourcing a staff member and a vessel to be on standby in case of rat incursion. They would then mobilise to deploy traps as soon as they were alerted to an incursion on any of the islands and deploy traps. If this work is carried out rapidly it is easier to remove the rats. If incursion response is slow then the rats will be more widely distributed and difficult to eliminate. If the RSPB incursion hub was up and running the Applicant could supply funding for the hub to continue following the end of the EU Biosecurity for LIFE Project.
- Unfortunately, the difficulties in quantifying the compensation benefits meant the incursion hub was not pursued as a compensation option. However, many conservation strategies prioritise maintaining sites that are in favourable condition over tackling sites that are very degraded and this measure is based on similar principles. Many SPAs for seabirds are managed, and without continued management would not stay in favourable condition. However, a measure aimed at maintaining current conditions does not result in any readily quantifiable benefit that can demonstrably offset the potential impacts of the wind farm.
6. TIER II: DIVERSIONARY FEEDING OF SPECIALIST PEREGRINE FALCON NESTS
6. TIER II: DIVERSIONARY FEEDING OF SPECIALIST PEREGRINE FALCON NESTS
6.1. Background
6.1. Background
- Kittiwakes and auks are vulnerable to predation from a range of avian predators including large gulls, raptors and corvids[33]. Eggs, chicks or even adults may be predated, and since seabirds are long-lived, the loss of even a few adults annually can become significant over time. Although predators are an important part of any healthy ecosystem, there are circumstances under which predation may be a cause for concern, and even of colony failure.
- Accordingly, although predation was generally considered to not be a significant general factor limiting seabird populations, specialist predators as a distinct and separate issue were discussed on several occasions during stakeholder consultation. Although many large gulls and raptors may consume the chicks of smaller species as a part of their diet, there are some individuals that develop specialisms for seabird chicks and may take little else during the period that the chicks are available. During the course of a breeding season these few individuals may take a surprisingly large number of chicks, which can even have a demonstrable impact on colony productivity (Smart & Amar 2018, Mason et al. 2021).
- The removal of a very small number of specialist predators can be a successful means of increasing the population of the prey species. For example, a study at Benidorm Island offshore of the western coast of Spain showed that the selective removal of only 16 Yellow-legged Gulls over the course of 3 years led to a 65% reduction in the number of European Storm-petrels killed. In turn, this subsequently increased survival rate by 16% and breeding success by 23% in the Storm-petrel population (Sanz-Aguillar et al. 2008).
- As ethical and environmental concerns over traditional forms of wildlife management increase, there is a trend toward selective management that involves tackling a few ‘problem individuals’. There is strong evidence of individual behaviours in wild animal populations, and there are now a growing number of studies that demonstrate the effectiveness of selective management in providing a mutually acceptable solution (Swan et al. 2017).
- However, the issues are complex: many predators themselves are protected species, they are also suffering population declines, and are of equal conservation importance to their prey. Whilst in theory it is possible that removing large gulls from the vicinity of Kittiwake nesting areas would very likely result in significant improvements in breeding success, this would not be palatable to conservation organisations on the basis that Herring Gull (Red) and Great Black-backed Gull[34] (Amber) are of conservation concern in their own right (Eaton et al. 2015, Lopez et al. 2022).
- On this basis, diversionary feeding (DF) was recommended as a means of potentially reducing predation pressure without resulting in adverse impacts on predator populations. DF involves regularly feeding the predator, reducing its need to forage and therefore reducing predation impacts on the prey species. The technique has been successfully used in a number of locations to reduce the impacts of raptors on other protected (or commercially important) avian species. For example, diversionary feeding of a pair of Common Kestrels Falco tinnunculus with white mice reduced the predation rate of Little Tern Sternula albifrons chicks at North Denes (Norfolk) by 88%, resulting in a doubling of Little Tern productivity in the years when diversionary feeding was undertaken (Smart & Amar 2018). Similarly, diversionary feeding of Red Kite Milvus milvus reduced predation of Northern Lapwing Vanellus vanellus, with productivity doubling on average in years when diversionary feeding was undertaken (Mason et al. 2021).
- Although DF may be viewed as ‘unnatural’, it is one of the few methods available that allows both predator and prey to successfully raise chicks without harm to either species. DF would only be required for a few weeks whilst the chicks of the prey species are present, allowing the predators to forage naturally for the remainder of the year. It is anticipated that DF would only be used to deal with a small number of specialist pairs or individuals, and on this basis would not be expected to cause an increase in predator populations.
- Feedback from NatureScot acknowledged that DF could work for Peregrine Falcon Falco peregrinus (if the nest site is accessible), although they point out that it is unclear how much predation of Kittiwake is due to Peregrines for the Forth sites and indicate that it would be difficult to calculate the level of effect DF might have without further evidence of the current impact of specialist predators.[35]
- Peregrines nesting around sea cliffs are known to take seabirds, with some individuals developing specialisms (Sutton & Loram 2021). Although the need to gather site specific data is acknowledged, it is conceptually it is feasible DF of specialist Peregrines to be progressed as a compensatory measure, although this would involve gathering field data during the 2023 breeding season.
- Of course, in many instances it may not be feasible to carry out DF: predator nest sites may be inaccessible, or access may entail disturbance of too many other birds nesting nearby, or nests may be too close together for DF to be effective. The objective of this work would be to positively identify situations where DF would be considered to have a high likelihood of success.
- The advantage of DF over many of the other suggested compensatory measures is that it involves a direct saving of adults (and/or chicks), as opposed to tackling other indirect factors (e.g. prey, human disturbance etc.) in order to bring about an improvement in productivity. It was suggested that diversionary feeding of other predator species could be trialled, although this suggestion was not popular. However, further information on this topic is presented in Section 7.2.
- Although NatureScot agreed that DF had some merit and acknowledged that it could potentially work successfully for Peregrine Falcon, some issues were raised. DF has not yet been trialled on this species, and it is unclear how much predation of Kittiwake is due to peregrines for the Forth sites. Therefore, a further evidence-gathering stage would be required to identify potentially accessible nest sites and establish what level of benefit DF may have. Since the concept would require further development, it was placed in Tier II as it is not deliverable as a compensatory measure without further investigatory work.
7. TIER III: OTHER MEASURES CONSIDERED
7. TIER III: OTHER MEASURES CONSIDERED
- Tier III measures were unpopular with stakeholders and for this reason were not taken forward. The reasons why measures were not progressed are stated in within the summary sections for each measure. They are not being pursued at the current time, as measures that were viewed positively have been progressed in preference.
7.2. Diversionary feeding of other avian specialist predators
7.2. Diversionary feeding of other avian specialist predators
- The concept of diversionary feeding is described in Section 6 in relation to Peregrine Falcon. Other specialist predators likely to be impacting on auks and Kittiwake in the local area include Great Black-backed Gull, Herring Gull and various corvid species (Carrion Crow Corvus corone, Eurasian Magpie Pica pica, Eurasian Jackdaw Corvus monedula and Common Raven Corvus corax). All of these species may act as specialist predators in some circumstances, although only Great Black-backed Gull would be capable of taking adult Kittiwakes and auks. Corvids primarily take eggs and young chicks. Herring Gulls would take a combination of eggs and chicks of all ages. Predation may be carried out by locally breeding birds or by predators that commute to the colony from elsewhere (see Hario et al. 1994).
- A literature review of the main predator species was undertaken (see below) to clarify what is understood about specialist predators and predator-prey relationships affecting the key species both in Firth of Forth area and further afield and to highlight areas where further research could be of benefit in progressing potential compensation though DF.
- Although this review focusses on DF, it is acknowledged that other techniques may also be worthy of further investigation, such as the use of canes, which have been successfully used to reduce large gull predation of breeding terns (Babcock & Booth 2020a).
7.2.2. Great Black-backed Gull
- Great Black-backed Gull is the only locally-breeding seabird able to take adult Puffins. They are also capable of eating adult Guillemots and Razorbills, although these are only taken occasionally. Great Black-backed Gulls also take large numbers of juvenile puffins (pufflings), as well as Guillemot, Razorbill and Kittiwake chicks. Some Great Black-backed Gulls are known to wait specifically to catch Puffins as they enter and leave burrows, especially if vegetation is dense (Harris & Wanless, 2011).
- A specialist pair of Great Black-backed Gulls would take in excess of 30 Puffin adults over the course of the breeding period. However, this figure is based on corpse searches carried out at the end of the breeding season on Craigleith and Isle of May[36] and will be an underestimate, as a proportion of birds will have already been consumed. If 3 Great Black-backed Gull pairs were fed (as was the case in the Red Kite and Kestrel studies), it could save a minimum of 90 adult Puffins, as well their dependent chicks (assuming that Puffins present will be part of a breeding pair). Specialist Great Black-backed Gull pairs also feed their chicks on pufflings (and rabbit), so assuming a predation rate of 3 Pufflings per Great Black-backed Gull nest per day and a 30 day chick period then 90 pufflings would be predated by a specialist pair over the course of a breeding season. On this basis feeding 3 specialist nests could (if successful) result in a saving of around 270 pufflings.
- Great Black-backed gulls also take eggs as well as adults and chicks. A study of Great Black-backed Gulls from Newfoundland showed that they exhibited a preference for Guillemot eggs, selecting them disproportionately to their abundance, and consuming 40% of Guillemot eggs laid at one site, Gull Island (Veitch et al. 2016). Work would be required to confirm whether this preference is exhibited locally, although it seems reasonable on the basis that Guillemots do not make nests but lay eggs directly onto ledges on cliffs, meaning that the eggs are more visible than those of the other species.
- Students from Aberdeen University have been investigating the impacts of Great Black-backed Gull predation of Puffins on Craigleith, following increases in Great Black-backed Gulls and declines in Puffins. In 2017, a corpse search at the end of the breeding season revealed over 130 dead Puffins[37]. In 2018, a repeat corpse search revealed 130 dead Puffins. Although most took small numbers only, one specialist pair had stockpiled 30 Puffin carcasses in a ‘midden’ (Figure 16, Forth Seabird Group 2019).
- There is currently a PhD student on the Isle of May researching the foraging ecology of Great Black-backed Gulls and their potential effects on Puffin populations. Results from tagging Great Black-backed Gulls showed that 3 out of 10 birds did not leave the island to forage offshore at all, instead relying entirely on auks (mostly Puffin) and rabbits. This study also showed that these specialist Great Black-backed Gulls nested on their own outside the main colonies (S. Lopez, pers. comm).
- Corpse searches carried out on the Isle of May in 2017 and 2021 revealed that 570 and 500 adult Puffins were taken by Great Black-backed Gulls during these years respectively[38]. Around 10% of the 79 pairs currently breeding on the Isle of May are believed to be Puffin specialists, although almost all Great Black-backed Gulls will take some Puffins and pufflings during the course of a breeding season. An earlier study when there were only 27 pairs of Great Black-backed Gulls on the Isle of May, showed that 3 specialist pairs were responsible for the death of 96 Puffins, half of the total number found during corpse searches (Finney 2002).
- Stable isotope analysis is currently being carried out on feather samples taken from Great Black-backed Gulls that have been ringed on the Isle of May, and it is hoped that there will be differences in signature between specialists and non-specialists. If so, it is possible that this technique can be used to confirm the prevalence of specialists within the colony. Current knowledge is based on pellets, which give an incomplete picture of colony behaviour (as some birds pellet in inaccessible areas). Stable isotope analyses of feathers would also help answer other questions about specialists, for example whether specialists are mostly male, and whether both members of a pair specialise. If the technique is successful, it could be employed at other colonies and for other species (e.g. Herring Gulls).
- Other unanswered questions include understanding why Great Black-backed Gulls choose to specialise, and whether feeding at a higher trophic level is more desirable than taking marine prey[39]. At present there does not seem to be any differences in productivity between specialists and non-specialists, although further years data are required to confirm this. It is also unclear whether another specialist will colonise if a specialist pair is removed, and whether the chicks of specialists become specialists themselves when they return to breed 5 years later.
- However, it is known that the proportion of Great Black-backed Gulls that hunt Puffins varies markedly between colonies, and therefore what is observed on the Isle of May might not be representative of the situation at other local SPAs. For example, on North Rona in the early 1970s there were ~2000 pairs of gulls and 6,000 pairs of Puffins and only 93 dead Puffins were found during corpse searches (Evans 1978). By contrast on Dun (St Kilda) the 30-40 pairs of Great Black-backed Gulls were all Puffin specialists and over a period of 4 years, these gulls killed at least 5,500 Puffins. It is of note that more Puffins were taken from an area where burrow density was low (5% compared to 1% of main colony), with the gulls selectively targeting immature birds (Harris 1980).
- A recent study analysing 1,035 pellets from Great Black-backed Gulls on Skokholm showed that overall Great Black-backed Gulls fed on seabirds with 48% taking mostly Manx Shearwater, 38% taking mostly Rabbits and 7% taking anthropogenic waste and 7% taking marine prey. Generalist pairs comprised 73% of the population with the remaining 27% being specialists. Of these specialist pairs, 5 were seabird specialists and 2 were mammal specialists. There were no links between diet and breeding success (Westerberg et al. 2019).
- Although specialist Great Black-backed Gulls have been identified on both on the Isle of May and Craigleith, it is possible that there may be specialists at other locations where Puffins and Great Black-backed Gulls nest in proximity such as Fidra, Inchkeith, Inchcolm Fowlsheugh, Longhaven and the Farne Islands. Although research has focussed on Puffins, the chicks of Kittiwakes, Guillemots and Razorbills will also be routinely taken by Great Black-backed Gull.
7.2.3. Herring Gull
- Herring Gulls predate the chicks and eggs of Puffins, Guillemots, Razorbills and Kittiwakes. They also impact on the auk species through kleptoparasitism[40]. Kleptoparasitism can be damaging because the chick may remain unfed for a significant amount of time (due to the long duration of many foraging bouts) at what is a critical growth period. In one part of the Isle of May on average 37% of Puffins bringing in fish were attacked and there was a negative relationship between Puffin recruitment and gull density (Harris 2011). However, looking at the Isle of May as a whole, only 7% of loads were lost, and there was no evidence that kleptoparasitism resulted in any detectable impact on reproductive output. At another site (Great Island), Puffins chose to nest in areas with high densities of Herring Gulls, perhaps because they gained some protection from Great Black-backed Gulls and Ravens by doing so (Pierotti 1983).
- It is known from various studies that Herring Gulls can specialise on other seabird species. For example, a study of Herring Gull predation of Lesser Black-backed Gull chicks carried out in the Gulf of Finland showed that two pairs of Herring Gulls were responsible for 17% of chick deaths. Once these predators were culled, predation rate reduced to 2% and fledging success improved significantly. However, ‘long-distance’ predation of chicks by Herring Gulls nesting elsewhere continued as these birds were hard to track and remove. This resulted in a decision to cull Herring Gulls by removing eggs from the entire Söderskär archipelago. During 4 years Herring Gull numbers were reduced by 40%, but this still did not stop the ‘long-distance’ predation. Although this study was carried out some years ago, it demonstrates the influence that specialist predators can exert (Hario et al. 1994).
- Another study of Herring Gulls in Newfoundland showed that 75–80% of Herring Gulls specialized on either intertidal organisms, human refuse, or other seabirds, while only 20–25% had generalized diets. Specific types of specialists tended to nest in particular habitats, and the study concluded that choice of nesting habitat may actually constrain diet choice. Whilst the intertidal specialists laid larger and heavier clutches, there were no significant differences in fledging rates, indicating the importance of other factors in fledging success as well as energy from prey (Pierotti & Annett 1991).
- A contrasting study of Herring Gull predation on Ring-billed Gulls Larus delawarensis nesting at Lake Ontario showed that predation was opportunistic, occurring primarily when Ring-billed Gulls entered the territory of Herring Gulls. Dietary studies revealed that nearly all Herring Gulls ate birds, but otherwise had a broadly similar diet, with birds accounting for a third of prey at most. These Herring Gulls could not be classified as bird specialists; predation of birds was generally infrequent, predation attempts were also often unsuccessful, and overall bird prey was considered insufficient to meet energetic requirements (Ingraham et al. 2020).
- Anecdotal evidence suggests that there may be Herring Gulls on the Isle of May that specialise on Kittiwake chicks: some individuals have been observed on multiple occasions watching Kittiwake nests, waiting for the moment that the adults swap over to then make their attack on the chick (F. Daunt, CEH, pers. comm.). A few Herring Gulls are also known to specialise in catching young Puffins while they are exercising their wings or when they are fledging (Harris 2011).
- There are a large number of Herring Gulls nesting alongside or in relatively close proximity to Kittiwakes and auks at a number of sites locally, including St Abbs to Fast Castle SPA (172 AON) Fowlsheugh SPA (1055 AON), and the Forth Islands SPA (5,964 AON) (JNCC 2021[41]). Within the Forth Islands SPA the Isle of May, Craigleith and Fidra support the largest Herring Gull colonies with 3061 AON, 1092 AON and 1059 AON respectively (Forth Seabird Group 2018 & 2019), with smaller numbers present on Fidra, Bass Rock, Inchcolm, Inchgarvie, Inchmickery, Inchkeith, Carr Craig, Haystack and Lamb. Small numbers are known to nest on Dunbar Castle, and also in Eyemouth town. There are likely to be other urban nesting locations in addition to these. Further investigation of the impacts of Herring Gulls on the key species at local colonies could clarify the likely scale of predation.
7.2.4. Other Predators
- Several corvid species may steal seabird eggs, such as Crows, Magpies, Jackdaws and Ravens. Some may also take chicks, as a camera trap study on North Ronaldsay reveals (Figure 7.2, taken from Johnston et al. 2019).
- A review of studies looking at predation as a limiting factor for bird populations concluded both that generalist predators such Crows (and Foxes) occur at high densities in the UK compared with other European countries, and that ground-nesting seabirds, waders and gamebirds can be limited by predation (Roos et al. 2018). In this study shearwaters, storm petrels, gulls, skuas, terns and auks were classified together as ‘seabirds’. After an experimental removal of predators, the majority of cases involving seabirds (80%) and gamebirds (81%) found an increase in the population size of the prey species, whereas this effect was smaller for other groups such as waders (45%), Passerines (40%) and Raptors and owls (33%). The paper concludes that in the long-term further investigation of land-use practices and landscape configurations should be undertaken to reduce predator numbers (Roos et al. 2018). Although these results are based on a review of multiple studies across a number of species groups, the results indicate that the impacts of corvid predation on seabirds is an area worthy of further study.
- Lesser Black-backed Gulls are generalist and opportunist feeders, known to take a range of marine prey, fisheries discards and human refuse (Mitchell et al. 2004). Even though they are classified as ‘generalist’ feeders, some individuals choose to specialise (Juvaste et al. 2017). Documented examples include specialisation on fisheries discards (Tyson et al. 2015), worms (Coulson & Coulson 2010) and human refuse (Juvaste et al. 2017). Although Lesser Black-backed gulls can and do take the chicks of other species, there is less evidence that individuals become seabird chick specialists.
- Great Skua it is a significant predator of Kittiwakes and auks. For example, on Foula (Shetland) Great Skua predation reduced the Kittiwake population by 54–85% between 1981-1995 (Heubeck et al 1999). It has been calculated that a 5% increase in seabirds in Great Skua diet at Foula is equivalent to consumption of an extra 1,000 Fulmars or 2,000 Kittiwakes (Votier et al. 2004a). A different study of Great Skua diet carried out on Hermaness (Shetland) showed that Great Skuas predated 12,500 and 13,000 seabirds in 1999 and 2001 respectively. In both years their prey was predominantly auks, with 7,837 taken in 1999 and 7,125 in 2001. Kittiwakes were also a significant prey item with 417 taken in 1999 and 285 in 2001 (Votier et al. 2004b). There are indications that rate of Great Skua predation on seabirds in some locations is unsustainable (Heubeck et al 1999, Regehr et al 1996), a situation that will have worsened following changes in fisheries policy and discard bans. Since there are no Great Skuas breeding within the Firth of Forth, Great Skua is not a problem locally. However, further investigation of Great Skua predation on auks and Kittiwakes elsewhere could be considered if further compensation is needed.
- Although this review has focussed on the species thought most likely to operate in this locality, sometimes unexpected specialist predators can occur. For example, in 2010 a Montagu’s Harrier Circus pygargus appeared at RSPB Minsmere (Suffolk) and stayed for two weeks during which period it took a large number of duck, wader and gull chicks.[42]
7.2.5. Summary
- There is reasonable evidence to suggest further investigation and in some instances trials of DF of other specialist avian predators could lead to success in reducing predation pressure on SPA populations of Kittiwake and auks. However, due to lack of support this work is not being taken forward.
- Although recent studies suggest that in many cases specialist Great Black-backed Gulls nest well away from the main colony, occupying their own territory in which they forage primarily on Puffins and rabbits (S. Lopez pers. comm.) and there is clear evidence to suggest that a small number of specialist Great Black-backed Gulls exert significant predation pressure on both adult Puffins and pufflings (as well as Razorbill, Guillemot and Kittiwake chicks), it was felt that DF of Great Black-backed Gull was not feasible on the basis that many birds nest colonially. Further investigation into the impacts of specialist Herring Gulls Larus fuscus on Kittiwake on the Isle of May (where the Herring Gull colony numbers 3,398 AON – The Forth Seabird Group 2019) was also rejected for the same reasons. There was a general reluctance to engage with measures to reduce predation pressure as a means of compensating for the potential impacts of the wind farm due to concerns regarding any undesirable or unforeseen impacts on predator populations. These concerns were not alleviated by suggestions that further study into the potential use of DF could be conducted as a research trial through a reputable academic institution.
7.3. Supplementary feeding of Puffin & Kittiwake
7.3. Supplementary feeding of Puffin & Kittiwake
7.3.1. Background
- The principle of supplementary feeding is to provide additional prey to chicks to support growth and development and ultimately colony productivity (if feeding is undertaken at sufficient scale). For this to work there must be a close link between prey supply and breeding success as is thought to be the case for Kittiwake (Carroll et al. 2017).
- Accordingly, the technique seems most likely to be successful when prey supply would otherwise become limiting and is perhaps of particular benefit to those species that lay several eggs and typically hatch more than one chick. In this situation the disadvantages experienced by the later hatching chick or chicks may theoretically be overcome with additional food provided that the prospects for survival are linked to prey supply and not driven by obligate siblicide for example.
- As a general principle, supplementary feeding is, of course, less desirable than measures directed towards improving the natural prey supply by a variety of means, or in the case of competing commercial fisheries, to leave ‘enough for the birds’ in the first place (Cury et al. 2011).
- Species that have a more restricted diet as specialists for one reason or another, may conceivably be more likely to benefit from than generalists, that may be able to switch to alternative prey relatively effectively should the supply of preferred prey become limiting. At least on the Isle of May and perhaps at other locations in and around the Firth of Forth, Puffin and Kittiwake are seen to be sandeel specialists (see Wanless et al. 2018) and it is envisaged that these species would benefit most from supplementary feeding. This is exacerbated by the fact that sandeels are prone to considerable fluctuation in abundance with an increasing prospect of poor recruitment as the climate warms promoting the prospect of mismatch between the timing of hatching of 0-group sandeels relative to their copepod prey.
- In fact, studies trialling supplementary feeding of both Puffin (Harris 1978) and Kittiwake (Gill et al. 2002) have been undertaken that show it can be an effective means of improving productivity especially if prey availability is limited and the adults are struggling to provision effectively. However, where prey supply is already adequate, supplementary provisions to a single chick in particular may have the effect of reducing the demands upon the parents, which in turn provision the chick less. In effect, there is no net gain in provisions for the chick and this may then become dependent on those provisions.
- Supplementary feeding of Kittiwake and Puffin during years of poor prey supply in particular has clear potential to be worthwhile. Whilst it is not desirable to feed birds regularly and certainly effort is better directed towards protecting prey offshore, it is considered that the decline of Kittiwake in particular is serious enough to warrant consideration of alternative techniques such as supplementary feeding that have not been trialled before in the UK.
- Although the concept has merit, and there is some scientific evidence to suggest it could be successful, a proof of concept trial would be required for Kittiwake to ensure that food could be delivered to nests without causing disturbance. However, ultimately the possibility of carrying out supplementary feeding was dismissed due to lack of stakeholder support. Nonetheless a description of how the work could be approached is included below.
7.3.2. REVIEW of MEASURE
- Supplementary feeding of Puffin chicks is technically straightforward as fish can be deposited directly into the burrow mimicking prey delivery by provisioning adults (Harris 1978). As a result, relatively large numbers of birds could conceivably be efficiently treated in a high-density locality such as the Isle of May once the logistics of a sufficient prey supply, such as frozen Sprat as used by Harris (1978) has been overcome.
- Supplementary feeding Kittiwake chicks is more complex as a result of their use of open nests sites, where neighbouring adults or other species (e.g. larger gulls or even corvids) may scavenge prey from them. In addition, chicks beg the returning adults for food, and it is this begging action that prompts the adult to regurgitate prey directly into the chicks gullet These complexities were overcome in an Alaskan study (Gill et al. 2002), although the study site is somewhat unique (discussed further below). For supplementary feeding to work well in the UK, the site would need to be selected carefully.
- Given the key differences between Puffin and Kittiwake, a burrow nesting species with a single chick and an open-nesting species with multiple chicks respectively, the two species are discussed separately below.
Supplementary feeding Puffin chicks
- Three studies involving supplementary feeding of Puffin chicks in Scotland were carried out some decades ago: one on St Kilda (Harris 1978) and two on the Isle of May (Harris 1978, Cook & Hamer 1997). Supplementary feeding has more recently been carried out in Norway on the Røst archipelago (Dahl 2005), where poor prey availability has led to a decrease in Puffin numbers of 81% between 1979–2019 (Fayet et al. 2021), including total breeding failure between 2007 and 2015.
- The study by Harris (1978), which trialled supplementary feeding on St Kilda and the Isle of May found that Puffin chicks at St Kilda fledged at significantly higher weights if they were provided daily with 50 g of sprats compared to control chicks (average weight of 316 g for 11 fed chicks vs. 301 g for 37 control chicks). Five chicks were removed from burrows and fed sprats ad libitum, and these chicks were even heavier (365 g), while three unfed chicks with a single parent were lighter (240 g). All removed chicks and fed chicks fledged, 37 of 39 controls fledged and three of six single-parent chicks fledged. There was no difference in fledging age between fed and control chicks (40 days), but single-parent chicks took longer to fledge (45 days). By contrast, Puffin chicks on the Isle of May exhibited smaller differences between treatments. Six chicks were removed and fed ad libitum, and these weighed on average 367 g. Ten chicks were fed with 50 g of sprats per day, and weighed 344 g. Control chicks weighed 331 g, whilst chicks of single parents weighed 303 g. Comparison of the data from St Kilda and the Isle of May suggests that the St. Kilda puffins were to some degree food limited, whereas those on the Isle of May were not at that time.
- A further supplementary feeding study on the Isle of May (Cook & Hamer 1997) was carried out to investigate whether there was a causal relationship between the nutritional status of pufflings and the subsequent rates of food provisioning by their parents. This study showed that chicks given supplementary food received less frequent meals from parents than control chicks, indicating that parents adjusted their foraging effort based on the nutritional status of the chick. If the growth rates of chicks were limited by poor food supply and adults delivered food as rapidly as they could, then regulation of food delivery would not be expected.
- Supplementary feeding of Puffin chicks on the Røst archipelago in Norway confirms that when chicks are fed parents reduce their foraging effort and the chicks do not receive any extra nutrition (Dahl et al. 2005). Whilst the study concluded that feeding conditions were adequate for normal breeding, further failures on the Røst archipelago indicate that prey is limiting and recent work at this site links large foraging ranges to lower chick provisioning rates (Fayet et al. 2021).
- Both of the studies carried out on the Isle of May discussed above were undertaken during a period when the Puffin colony was experiencing a period of rapid growth; conditions were considered to be favourable and chicks were rarely observed begging for food (Harris 1978). Figure 8.3 shows that since the latter Cook & Hamer (1997) study Puffin numbers have significantly declined, and it is anticipated that a further trial of supplementary feeding on the Isle of May will yield a different set of results.
Supplementary feeding Kittiwake chicks
- The supplementary feeding study at Middleton Island, Alaska (Gill et al. 2002) represents the only one yet conducted on Kittiwakes. In this study, both Kittiwake adults and chicks were fed on Herring ad libitum over two breeding seasons, and a range of parameters considered likely indicators of nutritional status were studied and ranked. Comparison of breeding performance were made between fed and unfed nests. Parameters considered included egg size, phenology, adult and chick behaviour, parental attendance, chick growth and survival and breeding success (Gill et al. 2002).
- Breeding success (specifically fledging success, hatching success and overall productivity), adult incubation and attendance during chick rearing and chick growth parameters (especially for the ß-chick[43]) were key variables that were positively influenced by supplemental feeding. Supplemental feeding improved fledging success of fed pairs by 6–8 standard deviations over unfed pairs, and hatching success was enhanced by 2.5–6 standard deviations (Gill et al. 2002).
- Kittiwakes typically lay either 1 or 2 eggs, although 3 is also possible. However, the latter is rarely recorded at colonies in the Firth of Forth for example. In clutches of more than 1 egg, laying occurs at about 2 day intervals, but sometimes there may be a delay to a third day. Hatching of eggs is asynchronous by about a day and occasionally longer. Single-egg clutches and the last egg in a 3-egg clutch have the lowest chance of producing a fledged chick. In 2-egg clutches, the first-laid egg is more successful than the second egg (Coulson 2011).
- There are reasons why the second chick (or ß-chick) is less successful: the first or α-chick learns to beg for food from the adult before the ß-chick hatches. To be successful the ß-chick has to be able to beg effectively enough to persuade the parents to feed it instead of its older sibling. As the second chick usually hatches a day or so after the first, this delay in feeding sometimes results in a greater difference in size than would be expected based on hatching date alone. If there is a third chick, it often dies, as the third egg is smaller and has a smaller yolk reserve, and it must also overcome the same challenges to get fed. However, the death should not be attributed to food shortage, but rather to the difficulty in being able to beg effectively enough to persuade the parents to feed it despite competition from the vigorous begging of its larger siblings (Coulson 2011).
- Supplemental feeding has been shown to improve productivity by improving the growth rate of the ß-chick. Growth rate has an important effect on subsequent post-fledging survival during the following two years. Productivity of single clutches is actually lower than that of larger clutches: of single-egg clutches 67% of the eggs failed to produce a fledged chick. (The rationale behind this is that higher quality females lay more eggs). Even when first time breeders are accounted for this is still poor. However, Coulson (2011) suggests that the problem lies with incubation, which in most cases finishes before hatching could occur. If the egg hatches, fledging success is the same as other clutches.
- Therefore, it seems reasonable to suggest that supplementary feeding not only improves productivity in conditions where prey is short, but even when prey is available it could also increase productivity by improving the growth rate (and therefore survival prospects) of the ß-chick. In the Alaskan study supplementary feeding also improved hatching success, meaning that more ß-chicks hatched, which in turn then benefitted from the supplementary feeding, resulting in an overall improvement in fledging success. (It is not known whether supplementary feeding would help the third or γ-chick, as none of the nests in the Alaskan study had 3 chicks. The egg of the γ-chick is smaller, so it may be that the γ-chick still dies even with supplementary food; a situation that requires further study. However given that the Kittiwakes rarely lay three eggs in the Firth of Forth then this is not considered a priority).
- A second supplementary feeding study of Kittiwakes from Middleton Island also demonstrates the indirect benefits of supplementary feeding (White et al. 2010). Siblicide may be observed in Kittiwake chicks, where the α-chick harasses one of the other chicks causing it to fall out of the nest as a consequence (Dickens 1987, 2021). The α-chick may also administer brief ‘punishing pecks’ to the b-chick to stop the latter from begging for food from a parent (Dickens 2021). White et al. (2010) provided supplemental food to Kittiwakes over a whole breeding season and compared the aggressive behaviour of fed chicks and control chicks. Control α-chicks showed more frequent and intense aggression than the fed α-chicks. The consequences of increased aggression for ß-chicks were lower begging rate, lower growth rate and lower survival rate. Thus, a sustained increase in food availability reduces broodmate aggression and improves ß-chick survival rates.
- It is also conceivable that supplementary feeding may encourage other birds to nest at the site resulting in a larger colony. This may have benefits, for example, in colony defence from potential predators.
- Although it seems likely that supplementary feeding would result in improved fledging success, there are some logistical issues. The Alaskan studies were carried out on Middleton Island where Kittiwakes nest on an abandoned radar tower that has been purposely adapted to carry out seabird research. The Kittiwakes nest behind removable panels, so that they can be fed, captured and weighed regularly (Figure 8.4).


- The trial site would need to be selected carefully as it would be difficult to carry out supplementary feeding at an open nest site where Herring Gulls and other scavengers would compete for the food. The site would need to be easily accessible and some means of depositing fish directly into the nest is required. Preliminary discussion with researchers suggested some form of tube system could be successful, with this retaining having a lid and with sufficient lubrication and/or some form of propulsion mechanism to ensure fish is delivered successfully. However, a portable rather than a fixed system is much preferred and at this stage, a very lengthy but still portable blow-pipe, may be the answer. Considering that technique development is required, a proof-of-concept trial would be needed in the first instance.
- The lack of stakeholder support and limited potential compensation benefits meant that this measure was not taken further.
7.4. Removal of plastic from the Firth of Forth
7.4. Removal of plastic from the Firth of Forth
7.4.1. Background
- Plastic pollution is now a universal issue for seabirds although the biological significance of ingestion is still unquantified at a population level. Even individual cases definitively attributing seabird mortality to plastic ingestion are rare (Pierce et al. 2004). This is because clinical data on the bird’s health is required when it is alive, followed by a necropsy. One documented example relates to an emaciated gannet brought into a wildlife rehabilitation centre, unable to stand or feed. It died shortly afterwards with necropsy confirming the cause of death was due to a blockage to the oesophagus caused by a bottle top. Patterns of ulceration in the gizzard show that the bottle top had also previously been lodged there too (Pierce et al. 2004). Ingestion of plastics in general is likely to be exacerbated by poor prey availability as hunger reduces prey discrimination.
- Plastic may impact on seabirds in various ways: it may be ingested resulting in a range of lethal and sub-lethal effects or incorporated into nests where it may cause entanglement (Votier et al. 2011, Roman et al. 2020). Physical impacts of plastic ingestion include suppression of appetite and lack of energy, which may impact on ability to forage and reproduce. In severe cases ingestion may causes blockages and death (Pierce et al. 2004, Roman et al. 2020). Plastic ingestion also has chemical effects, concerning the transfer of plastic-additive and plastic- adsorbed chemicals (Roman et al. 2020).
- CEH regularly collect dead Puffins found on the Isle of May to monitor them, and a routine dissection showed that many contain nurdles (small industrial pellets used to make plastic). It is considered highly likely that Guillemot, Razorbill and Kittiwake will also contain high levels of nurdles, especially Kittiwake as they pick small items off the surface (F. Daunt CEH, pers. comm). Nurdles resemble fish eggs, making them attractive to many marine species. Persistent organic pollutants from the surrounding seawater adsorb onto the surface of nurdles, concentrating toxins to levels millions of times higher than the surrounding water (Mato et al. 2001). Beaches around the Firth of Forth, in particular Queensferry, are known to have high numbers of nurdles[44].
- Understanding that prey abundance is low and that plastic pollution is a problem within the Firth of Forth, then reducing marine plastics is intuitively beneficial both for seabirds and other marine life, although quantifying the benefits is unfortunately not possible. Whilst nurdles cannot yet be removed from the marine environment, user plastic certainly can be, and relatively straightforward solutions are available. Beach cleans and regulation of rubbish entering watercourses are effective. Marine litter can also be removed by other methods such as clean-up vessels (purpose designed vessels to clean-up harbours) and through novel technologies currently under development by companies such as Icthion, including filters that can be retrofitted to shipping vessels to collect plastic without impacting on marine life. Although fitting filters to vessels could not be viewed as compensation per se, it is possible that these kinds of initiatives could be incorporated into elements of the project further down the line.
- The possibility of initiating further work on nurdles and seabirds within the Firth of Forth was discussed with Fidra, an East Lothian based charity that has pioneered work on nurdles and which runs The Great Nurdle Hunt, an initiative that encourages the public to look for nurdles and submit data in order to improve awareness and target potentially polluting industries. However, Fidra do not work directly on seabirds, and on this basis the possibility was discounted.
- Beach cleans and regulation of rubbish entering water courses was pursued instead. One possibility identified was re-invigoration of the Forth Estuary Forum’s Coastal Litter Campaign (Storrier et al. 2004), which ran for three years between 2001 and 2004, and made substantial progress towards a ‘litter-free Forth’. The project used a team of volunteers, who used a scientifically standardised technique to assess trends in the deposition of fresh marine litter each month. The campaign found that the majority of the litter came from land-based sources, which became the focus of their campaigns. However, The Forth Estuary Forum did not respond to our invitation to discuss this further in time for the compensation scoping process, although a reply was received some months later indicating that a collaboration to tackle plastic pollution in The Forth Estuary would be of interest.
- Stakeholder consultation relating to potential rodent eradication from Inchcolm revealed minor issues with plastic pollution, and as a result removal of plastic litter from beaches was incorporated into work proposed at this site (see Section 2.3).
- Ingestion of plastics in general is likely to be exacerbated by poor prey availability as hunger reduces prey discrimination. Understanding that prey abundance is low and plastic pollution is a problem within the Firth of Forth, then reducing marine plastics is intuitively beneficial both for seabirds, seals and other marine life, although quantifying the benefits is unfortunately not possible.
- Feedback from NatureScot indicated that whilst they agreed that removing plastics would have positive effects, they did not consider that it could be classified as a compensatory measure because the benefit is impossible to quantify[45]. On this ground further work on reducing the impacts of plastic pollution on seabirds has not been pursued.
7.5. Fox control at Badbea and/or Longhaven cliffs
7.5. Fox control at Badbea and/or Longhaven cliffs
7.5.1. Background
- During stakeholder consultation it was raised that foxes had started to make their way onto the boulder beach at Badbea Cliffs, and as a consequence the Shag colony failed. Guillemot and Razorbill nesting in this area were also affected. Foxes were also identified as a problem at Longhaven Cliffs. Locally, Foxes are believed to have increased following the ban on hunting, and it is likely that other mainland sites certainly within North and East Caithness SPAs and possibly in other mainland locations will be affected too general, Fox control is likely to result in the greatest benefit for Puffin and Razorbills as cliff-nesting birds would not be vulnerable.
- Further discussion with SWT in relation to foxes at Longhaven Cliffs indicated that although foxes are an issue, control would be difficult at this location and would require sustained stakeholder consultation with multiple landowners as only a small amount of land adjacent to the cliffs is owned by SWT.
- Due to the nature of the habitat, particularly at Longhaven, it may be difficult to eliminate foxes as it is inevitable that territories will be colonised by new animals. The possibility of a fox fence at Longhaven was discussed, but this was not considered feasible as it would significantly disrupt the aesthetic of the reserve for what is a questionable benefit.
- The issue of a fox/foxes at Badbea was more surprising as the site is difficult to access. The fox managed to descend the cliff and transverse a boulder field to access the nests. This was considered an unexpected achievement for a fox, and if the individual or group of animals involved were removed, other foxes would not necessarily repeat this behaviour. Guillemot, Razorbill, Shag and Herring Gull nest in the area suffering from fox predation.
- Lethal fox control is readily undertaken by experienced, licenced professionals. Control is typically required on an annual basis and monitoring is valuable to evaluate the effect. This becomes particularly meaningful when established over a long timescale.
- The scale of the benefit of such measures are difficult to judge but could run into hundreds of eggs, chicks and adults annually and if prevented could result in greater productivity. However, without any indication of the number of foxes that might be present, there is uncertainty relating to the current effect of predation. Since fox control is lethal, it was felt that this would not be viewed favourably, especially in view of negativity regarding less impactful methods of reducing predation such as DF.
8. Conclusion
8. Conclusion
- Two colony-based measures are proposed as compensatory measures for the Proposed Development and several others have been explored within this report as part of a thorough compensatory measure identification and selection process. It is proposed that the final measures to take forward are:
- Rat eradication and biosecurity to benefit Kittiwake, Guillemot, Razorbill and Puffin nesting at Handa Island;
- Safeguarding the Dunbar Kittiwake colony through wardening and targeted work to reduce human disturbance and other colony-related pressures;
- The strengths and weaknesses of all the measures are summarised in Table 8.1. In the case of the two Tier I measures proposed the strengths clearly outweighed the weaknesses.
- The strengths of removing rats from Handa exceeded the weaknesses. In this case the key strengths included large compensation benefits (see Table 8.2), and the provision of the required resource to maintain biosecurity at Handa in the long term, whilst the only weakness was the distance of Handa from the Firth of Forth. Although a search was conducted to find a closer compensation site (see Section 2.2), none were comparable in terms of compensation benefits.
- With regards to Dunbar, the main strengths included proximity to and connectedness with colonies within the Forth Islands SPA, and also the evidence regarding the role of human disturbance in restricting birds from key nesting areas. The key weakness identified was that it may not be possible to tackle all of the disturbance sources effectively. However, this would be a very poor reason not to undertake the work as there is much that can be readily achieved at Dunbar to improve habitat quality for the Kittiwakes (see Section 3).
- With regards to the Tier II measures, although both the incursion hub and diversionary feeding of Peregrine Falcon were of clear benefit, there were more significant weaknesses. With regards to the incursion hub, the impossibility of quantifying the benefits caused significant difficulties, whereas the need for a field trial meant that diversionary feeding was less attractive than other methods that could be implemented more rapidly.
- The Tier III measures were largely unpopular with stakeholders and on this basis were considered weak. Of the Tier III measures, removal of plastics proved impossible to quantify in terms of likely compensation benefits and was placed in Tier III in response to feedback from NatureScot who advised they would not considered it as a compensatory measure. Diversionary feeding of other avian predators (i.e. Great Black-backed Gull) and supplementary feeding of Kittiwake and/or Puffin were also unpopular with stakeholders (as well as being unproven) and similarly were placed in Tier III. Based on other feedback from stakeholders on measures that involved control of predators, it was anticipated that lethal control of foxes at Badbea would similarly be unpopular. Since compensatory measures need to have support, these weaknesses were viewed to outweigh any potential strengths.
- The compensation benefits for each of the Tier I measures which are proposed as compensatory measures, expressed as Conservation Targets, are summarised in Table 8.2. Although in reality compensation benefits would not be delivered annually in this manner, the numbers are presented in this metric in order to enable comparison with the modelled annual mortality from the wind farm.
Table 8.1: Colony-based compensatory measures and their prioritisation ranking.
9. References
9. References
ABPmer (2022). Identification of Seabird Islands for Rodent Removal. Seabird Compensation: Berwick Bank Offshore Wind Farm. Report No. R3932. Report prepared for SSE Renewables.
Allbrook, D. & Quinn, J. (2020). The effectiveness of regulatory signs in controlling human behaviour and Northern Gannet (Morus bassanus) disturbance during breeding: an experimental test. Journal for Nature Conservation 58, 125915.
Amaral, J., Almeida, S., Sequira, M., & Neves, V. (2010). Black rat Rattus rattus eradication by trapping allows recovery of breeding roseate tern sterna dougalii and common tern S. hirundo populations on Feno Islet, the Azores, Portugal. Conservation Evidence 7, 16-20. Available online: Paper1491BlackRatEradicationTernsAzores[2010,7,16-20] (conservationevidencejournal.com)
Aragundi, S. (1994). The effects of the predation by the brown rat on Fulmars on Handa Island. Unpublished MSc thesis, Aberdeen University.
Araújo, M.S., Bolnick, D.I. & Layman, C.A. (2011). The ecological causes of individual specialisation. Ecology Letters 14: 948-958.
Arnott, S.A. & Ruxton, G.D. (2002). Sandeel recruitment in the North Sea: demographic, climatic and trophic effects. Marine Ecology Progress Series, 238, 199-210.
Arroyo, B. & Razin, M. (2006) Effect of human activities on bearded vulture behaviour and breeding success in the French pyrenees. Biological Conservation, 128, 276-284.
Babcock, M. & Booth, V. (2020a). Tern Conservation Best Practice: Using Canes to Deter Avian Predators. LIFE13 NAT/UK/000384.
Babcock, M. & Booth, V. (2020b). Tern Conservation Best Practice. Anti-predator Fencing. LIFE14 NAT/UK/000394.
Babcock, M., & Booth, V. (2020c). Tern Conservation Best Practice: Diversionary feeding kestrels. Supported by EU LIFE project ‘Improving the conservation prospects of the priority species roseate tern throughout its range in the UK and Ireland’ LIFE14 NAT/UK/000394.
Banks, A.N., Coombes, R.H. & Crick, H.Q.P. (2003). The Peregrine Falcon breeding population of the UK and Isle of Man in 2002. A report to the British Trust for Ornithology, Raptor Study Groups and to the Statutory Conservation Agencies/RSPB Annual Breeding. Birds Scheme.
Barton, N.W.H. & Houston, D. C. (1993). A comparison of digestive efficiency in birds of prey. Ibis 135: 363–371.
Beale, C.M., Monaghan, P. (2004). Human disturbance: people as predation-free predators? Journal of Applied Ecology 41:335-343.
Bell, P. (2014). Predator Free Rakiura Halfmoon Bay Project – analysis of options for proposed predator fence. Discussion document prepared by the Department of Conservation for the Predator Free Rakiura (PFR) Governance Group, c/o Southland District Council, Invercargill 17 p.
Birkhead, T.R. (1977). The effect of habitat and density on breeding success in the Common Guillemot (Uria aalge), Journal of Animal Ecology, 46: 751-764.
Boag, D., & Alexander, M. (1986). The Atlantic Puffin. Blanford Press, Poole.
Bolduc, F. & Guillemette, M. (2003) Human disturbance and nesting success of Common Eiders: interaction between visitors and gulls. Biological Conservation, 110 77-83.
Bolnick, D., Amarasekare, P., Araújo, M.S., Bürger, R., Levine, J.M., Novak, M., Rudolf, V.H., Schreiber, S.J., Urban, M.C. & Vasseur, D.A. (2011). Why intraspecific trait variation matters in community ecology. Trends in Ecology & Evolution, 26:183-192.
Bolton, M., Conolly, G., Carroll, M., Wakefield, E.D. & Caldow, R. (2018). A review of the occurrence of inter-colony segregation of seabird foraging areas and the implications for marine environmental impact assessment. Ibis. Doi.10.1111/ibi.12677.
Bötsch, Y., Tablado, Z., Scherl, D., Kéry, M., Graf, R.F., Jenni, L. (2018). Effects of recreational trails on forest birds: human presence matters. Frontiers in Ecology and Evolution, https://doi.org/10.3389/fevo.2018.00175.
Bradbury, G., Shackshaft, M., Scott-Hayward, L., Rextad, E., Miller, D. & Edwards, D. (2017). Risk assessment of seabird bycatch in UK waters. Report to Defra. Defra Project: MB0126.
Brooker, H., Slader, P., Price, D., Bellamy, A.J., & Frayling, T. (2018). Cliff nesting seabirds on Lundy: Population Trends from 1981 to 2017. Journal of the Field Society of Lundy 6: 65-76.
Bull, J., Wanless, S., Elston, D.A., Daunt, F., Lewis, S. & Harris, M.P. (2004). Local-scale variability in the diet of Black-legged Kittiwakes Rissa tridactyla. Ardea 92(1): 43-52.
Cain, I. et al (2022). Berwick Bank Wind Farm: Inchcolm Predator Eradication Feasibility Study. Contract report prepared for SSE Renewables.
Calado, J., Ramos, J., Almeida, A., Oliveira, N. & Paiva, V. (2020). Seabird-fishery interactions and bycatch at multiple gears in the Atlantic Iberian coast. Ocean & Coastal Management 200:105306 doi:10.1016/j.ocecoaman.2020.105306.
Caravaggi, A., Cuthbert, R.J., Ryan, P.G., Cooper, J. & Bond, A.L. (2018). The impacts of introduced House Mice on the breeding success of nesting seabird on Gough Island. Ibis. doi.10.1111/ibi.12664.
Carroll, M.J., Bolton, M., Owen, E., Anderson, G.Q.A., Mackley, E.K., Dunn, E.K. & Furness, R.W. (2017). Kittiwake breeding success in the southern North Sea correlates with prior sandeel fishing mortality. Aquatic Conservation: Marine & Freshwater Ecosystems 27: 1164–1175.
Coleman, J.T., Coleman, A.E., Rickeard, A. & Anderson, R. (2011). Long-term monitoring of a colony of Black-legged Kittiwakes Rissa tridactyla in Scotland. Ringing and Migration 26, 9-14.
Collins, P.M., Green, J.A., Dodd, S., Shaw, P.J.A. & Halsey, L.G. (2014). Predation of Black-legged Kittiwake Chicks Rissa tridactyla by a Peregrine Falco peregrinus: Insights from time-lapse Cameras. The Wilson Journal of Ornithology 126(1):158-161.
Comhairle nan Eilean Siar (Western Isles Council) Development Department. Socio Economic Update No. 37, May 2018.
Cook, M.I., & Hamer, K.C. (1997). Effects of supplementary feeding on provisioning and growth rates of nestling Puffins Fratercula arctica: evidence for regulation of growth. Journal of Avian Biology, Vol 28, No. 1 pp. 56-62.
Cortés-Avizanda, A., Carrete M., Serrano, D., Donázar, J.A. (2009). Carcasses increase the probability of predation of ground-nesting birds: a caveat regarding the conservation value of vulture restaurants. Animal Conservation 12:85-88.
Coulson, J.C. (1963). The status of the Kittiwake in the British Isles. Bird Study, 10, 147-167.
Coulson, J.C. (1982). The changing status of the Kittiwake Rissa tridactyla in the British Isles 1969-1979. Bird Study 30, 8-16.
Coulson, J.C. (2011). The Kittiwake. T & AD Poyser, London, UK. ISBN: 978-1-4081-0966-3.
Coulson, J.C. & Coulson, B.A. (2008). Lesser Black-backed Gulls Larus fuscus nesting in an inland colony: the importance of earthworks (Lumbricidae) in their diet. Bird Study, 55, 3: 297-303.
Coulson, J.C. & White, E. (1959). The post-fledging mortality of the Kittiwake. Bird Study, 6:3, 97-102.
Cramp. S., Bourne, W.R.P., & Saunders, D. (1974). The Seabirds of Britain and Ireland. Collins, London.
Carney, K.M. & Sydeman, W.J. (1999). A review of Human Disturbance Effects on Nesting Colonial Waterbirds. The International Journal of Waterbird Biology 22(1): 68-79.
Coleman, J.T., Coleman, A.E., Rickeard, A., & Anderson, A. (2011). Long-term monitoring of a colony of Black-legged Kittiwakes Rissa tridactyla in Scotland. Ringing and Migration 26, 9-14.
Cramp, S., Bourne, W.R.P., & Saunders, D. (1974). The Seabirds of Britain and Ireland. Collins, London, UK.
Cury, P.M., Boyd, I. L., Bonhommeau, S., Anker-Nilssen, T., Crawford, R. J. M., Furness, R.W., Mills, J.A. Murphy, E.J., Österblom, H., Paleczny, M., Piatt, J.F., Roux, J.-P., Shannon, L. & Sydeman, W.J. (2011). Global seabird response to forage fish depletion—one-third for the birds. Science 334: 1703–1706.
Dahl, H.K, Barrett, R.T., & Ims, R.A. (2005). Effects of supplementary feeding on provisioning and growth rates of Atlantic Puffin Fratercula arctica chicks in north Norway. Atlantic Seabirds 7(3): 133-143.
Day, M. (Director) (2010). The Guga Hunters of Ness. BBC Documentary produced by Intrepid Cinema.
De Pascalis, F., Collins, P.M., & Green, J.A. (2018). Utility of time-lapse photography in studies of seabird ecology. PLoS ONE 13(12): e0208995. https://doi.org/10.1371/journal.pone.0208995.
Dickens, D.W. & Clark, R.A. (1987). Games theory and siblicide in the kittiwake gull, Rissa tridactyla. Journal of Theoretical Biology 125:3, 301-305.
Dickens, D.W. (2021). “Punishing” pecks and “siblicidal” pecks in kittiwake (Rissa tridactyla) chicks. Learning and motivation. https://doi.org/10.1016/j.lmot.2020.101695.
DTA Ecology (2021). Framework to evaluate ornithological compensatory measures for offshore wind. Process Guidance Note for Developers. Doc. Ref. 1107 Marine Scotland CM framework.
Eaton M.A., Aebischer N.J., Brown A.F., Hearn R.D., Lock L., Musgrove A.J., Noble D.G., Stroud D.A. & Gregory R.D. (2015). Birds of Conservation Concern 4: the population status of birds in the United Kingdom, Channel Islands and Isle of Man. British Birds 108: 708–746.
ECON & ABPmer (2022). Berwick Bank. Evidence and recommendations for compensatory measures to support a Habitats Regulations Derogation. Internal report prepared for SSER.
Edinburgh Natural History Society Report (1992). Forth Island Seabird Counts 1992. Available online at: http://www.forthseabirdgroup.org.uk/pages/Reports.htm
Evans, P.G.H. (1978). The birds of North Rhona and Sula Sgeir. Hebridean Naturalist 1978: 21-36.
Fayet, A., Clucas, G.V., Anker-Nilssen, T., Syposz, M., & Hansen, E.S. (2021). Local prey shortages drive foraging costs and breeding success in a declining seabird, the Atlantic puffin. Journal of Animal Ecology doi: 10.1111/1365-2656.13442.
Fife Seal Group 2020. Annual Report 2020.
Finney, S.K. (2002). The dynamics of gull-puffin interactions: implications for management. PhD thesis, University of Glasgow.
Forth Estuary Forum. Forth Sight Newsletter, Issue No. 27, Spring 2015. https://www.forthestuaryforum.co.uk/wp-content/uploads/27-Newsletter-final.pdf
Forth Islands Heritage Group (2020). Annual Report 2020.
Forth Islands Heritage Group (2021). Annual Report 2021. Part 1.
Forth Seabird Group (2014). Forth Island Bird Report 2014. Accessible online at: http://www.forthseabirdgroup.org.uk/pages/Reports/FSG_Report_2014.pdf
Forth Seabird Group (2018). Forth Islands Bird Report 2018.
Forth Seabird Group (2019). Forth Islands Bird Report 2019.
Fredericksen, M., Wanless, S., Harris, M.P., Rothery, P. & Wilson, L.J. (2004). The role of industrial fishery and oceanographic change in the decline of North Sea black-legged Kittiwakes. Journal of Applied Ecology 41: 1129-1139.
Furness, R.W., MacArthur, D., Trinder, M. & MacArthur, K. (2013). Evidence review to support the identification of potential conservation measures for selected species of seabirds. Report to Defra. 158 pp.
Gabrielsen, G.W., Mehlum, F & Nagy, K.A. (1987). Daily energy expenditure and energy utilization of free-ranging Black-legged Kittiwakes. Condor 89:126-132.
Gabreilsen, G.W., Klaassen, M. & Mehlum, F. (1992). Energetics of Black-legged Kittiwake Rissa tridactyla chicks. Ardea 80:29-40.
Gibb, F.M., Régnier, T., Donald, K., & Wright, P.J. (2017). Connectivity in the early life history of sandeel inferred from otolith microchemistry. Journal of Sea Research 119:8-16.
Gilbert, G., Gibbons, D.W., & Evans, J. (1998). Bird Monitoring Methods. A manual of techniques for key UK species. Spectrum Flair Press, UK. ISBN 1 901930033.
Gill, J. A. (1996) Habitat choice in wintering pink-footed geese:quantifying the constraints determining winter site use. Journal of Applied Ecology, 33, 884-892
Gill, J.A. (2007). Approaches to measuring the effects of human disturbance on birds. Ibis https://doi.org/10.1111/j.1474-919X.2007.00642.x
Gill, V.A. 1999. Breeding performance of Black-legged Kittiwakes (Rissa Tridactyla) in relation to food availability: a controlled feeding experiment. MSc Thesis, University of Alaska, Anchorage.
Gill, V.A., Hatch, S.A. & Lanctot, R.B. (2002). Sensitivity of breeding parameters to food supply in Black-legged Kittiwakes Rissa tridactyla. Ibis 144: 268–283.
GoBe Consultants & Ørsted (2020). Response to the Secretary of State’s Minded to Approve Letter, Appendix 2: Kittiwake Compensation Plan. September 2020. pp. 28.
Grémillet, D., Péron, C., Provost, P. & Lescroël, A. (2015). Adult and juvenile European seabirds at risk from marine plundering off West Africa. Biological Conservation 182: 143–147.
Grémillet, D., Péron, C., Lescroël, A., Fort, J., Patrick, S.C., Besnard, A. & Provost, P. (2020). No way home: collapse in northern gannet survival rates point to critical marine ecosystem perturbation. Marine Biology 167: 189 doi.org/10.1007/s00227-020-03801-y.
Gummer, H., (2003), Chick translocation as a method of establishing new surface-nesting seabird colonies: a review, DOC Science Internal Series 150, Department of Conservation, Wellington.
Hamer, K.C., Phillips, R.A., Wanless, S., Harris, M.P., & Wood, A.G. (2000). Foraging ranges, diets and feeding locations of gannets Morus bassanus in the North Sea: evidence from satellite telemetry. Marine Ecology Progress Series 200: 257-264.
Hamer, K.C., Humphreys, E.M., Garthe, S., Hennicke, J., Peters, G., Grémillet, D., Phillips, R.A., Harris, M.P. & Wanless, S. (2007). Annual variation in diets, feeding locations and foraging behaviour of gannets in the North Sea: flexibility, consistency and constraint. Marine Ecology Progress Series 338: 295–305.
Hario, M. (1994). Reproductive performance of the nominate Lesser Black-backed Gull under the pressure of Herring Gull predation. Ornis Fennica 71:1-10.
Harris, M.P. (1978). Supplementary feeding of young puffins Fratercula arctica. Journal of Animal Ecology 47: 15–23.
Harris, M.P. (1980). Breeding performance of puffins Fratercula arctica in relation to nest density, laying date and year. Ibis 122:193-209.
Harris, M.P. (1996). Philopatry in the Common Guillemot Uria aalge. Bird Study, 43 (2) p. 134-137.
Harris, M.P. & Rothery, P. (1988). Monitoring of Puffin burrows on Dun, St Kilda, 1977-1987. Bird Study DOI:10.1080/00063658809480385
Harris, M.P., Albon, S.D. & Wanless, S. (2016). Age-related effects on breeding phenology and success of Common Guillemots Uria aalge at a North Sea colony. Bird Study, 63, pp. 311-318.
Harris, M.P. and Birkhead, T.R. (1985). Breeding ecology of the Atlantic Alcidae. In D.N. Nettleship and T.R. Birkhead (eds.). The Atlantic Alcidae. Academic Press, London.
Harris, M.P. and Wanless, S., (1987). The breeding biology of Guillemots Uria aalge on the Isle of May over a six-year period. Ibis, 130: 172-192
Harris, M.P. & Wanless, S. (2011). The Puffin. T & AD Poyser, London, UK. ISBN: 978-1-4181-0867-3.
Harris D, Gregory SD, Bull L, Courchamp F (2012) Island prioritization for invasive rodent eradications with an emphasis on reinvasion risk. Biol Invasions 14:1251–1263.
Harris, M.P., Albon, S.D. & Wanless, S. (2016). Age-related effects on breeding phenology and success of Common Guillemots Uria aalge at a North Sea colony. Bird Study https://doi.org/10.1080/00063657.2016.1202889
Harvey-Brown, J.A., & Buckley, T.E. (1887). A Vertebrate Fauna of Sutherland, Caithness and West Cromarty pp.38-40. David Douglas, Edinburgh.
Harvey-Brown, J.A. & MacPherson. H.A. (1904). A fauna of the North West Highlands and Skye pp. xliii-lii and 356-7. David Douglas, Edinburgh.
Heubeck, M., Mellor, R.M., Harvey, P.V., Mainwood, A.R. & Riddington, R. (1999). Estimating the population size and rate of decline in Kittiwakes Rissa tridactyla breeding in Shetland 1981-97. Bird Study 46, 48-61.
Hipfner, J.M. and Dussureault, J. (2001). The occurrence, size, and composition of Razorbill Nest Structures. The Wilson Bulletin, 113: 445-448.
Horswill, C. & Robinson R. A. 2015. Review of seabird demographic rates and density dependence. JNCC Report No. 552. Joint Nature Conservation Committee, Peterborough.
Ingraham, L.R., Oswald, S.A., Son, E., & Arnold, J.M. (2020). Opportunistic predation of birds by breeding Herring Gulls (Larus argentatus). PLoS ONE 15(10): e0239180. https://doi.org/10.1371/journal.pone.0239180
Kaiser, M. J., Galanidi, M., Showler, D. A., Elliott, A. J., Caldow, R. W. G., Rees, E. I. S.,Stillman, R. A. & Sutherland, W. J. (2006) Distribution and behaviour of Common Scoter Melanitta nigra relative to prey resources and environmental parameters. Ibis,148, 110-128.
Kaplan, B.S., O’Riain, M.J., van Eeden, R., & King, A.J (2011). A low-cost manipulation of food resources reduces spatial overlap between baboons (Papio ursinus) and humans in conflict. International Journal of Primatology 32:1397-1412.
King, C.M. & Forsyth, D.M. (eds.) (2021). The Handbook of New Zealand Mammals (3rd Ed). Oxford University Press. Auckland, New Zealand and CSIRO Publishing, Australia. 576 p.
Kubasiewicz, L.M, Bunnefeld, N., Tulloch, A.I.T., Quine, C.P. & Park, K.J. (2016). Diversionary feeding: an effective management strategy for conservation conflict? Biodiversity Conservation 25:1-22.
JNCC (Joint Nature Conservation Committee) (2021). Seabird Population Trends and Causes of Change: 1986–2019 Report (https://jncc.gov.uk/our-work/smp-report-1986-2019). Joint Nature Conservation Committee, Peterborough. Updated 20 May 2020.
JNCC (2021). Seabird Population Trends and Causes of Change: 1986-2019 Report (https://jncc.gov.uk/our-workshop/smp-report-1986-2019). Joint Nature Conservation Committee, Peterborough. Updated 20 May 2021. Available online at: SMP Report 1986–2019 | JNCC - Adviser to Government on Nature Conservation
Johnston, D.T., Furness, R.W., Robbins. A.M.C., Tyler, G., & Masden, E.A. (2019). Camera traps reveal predators of breeding Black Guillemots Cepphus grille. Seabird 32: 72-83.
Jones, T., Smith C., Williams, E. & Ramsay, A. (2008). Breeding performance and diet of Great Skuas Stercorarius skua and Parasitic Jaegers (Arctic Skua) S. parasiticus on the west coast of Scotland. Bird Study, 55, 257-266.
Juvaste, R., Arriero, E., Gagliardo, A., Holland, R., Huttunen, M.J., Muller, I., Thorup, K., Wikelski, M., Hannila, J., Penttinen, M-L., & Wistbacka, R. (2017). Satellite tracking of red-listed nominate Lesser Black-backed Gull (Larus f. fuscus): Habitat specialisation in foraging movements raises novel conservation needs. Global Ecology and Conservation 10: 220-230.
Lavers, J., Jones, I.L., & Diamond, A.W. (2007). Natal and breeding dispersal of Razorbills (Alca torda) in Eastern North America. Waterbirds 30(4): 588-593.
Lavers, J.L., Jones, I.L. & Diamond, A.W. Age at first return and breeding of Razorbills (Alca torda) on the Gannet Islands, Labrador and Machias Sea Island, New Brunswick, Waterbirds, 31, (1), pp.30-34. ISSN 1524-4695/.
Lavers, J.L., Jones, I.L. & Diamond. A.W. (2008). Age at first return and breeding of Razorbill (Alca torda) on the Gannet Islands, Labrador and Machias Seal Island, New Brunswick. Waterbird Biology 31, No. 1, pp.30-34.
Liley, D. (1999) Predicting the consequences of human disturbance, predation and sea-level rise for Ringed Plover population size. School of Biological Sciences. University of East Anglia, Norwich.
Liley, D., & Panter, C. (2017). First year results of monitoring bird disturbance around the Solent, in the presence/absence of rangers. Report prepared by Footprint Ecology for the Solent Bird Aware Initiative.
Liley, D. & Sutherland, W. J. (2007) Predicting the population consequences of human disturbance for Ringed Plovers Charadrius hiaticula: a game theory approach. Ibis, 149, 82-94
LIFE 13 NAT/UK/000209. Protecting and restoring the Shiant Isles SPA through rat removal, and safeguarding other seabird island SPAs in the UK.
Lopez, S.L., Bond, A., O’Hanlon, N., Wilson, J., Vitz, A., Mostello, C.S., Hamilton, F., Rail, J.F., Welch, L., Boettcher, R., Wilhelm, S.I., Anker-Nilssen, T., Daunt, F. & Masden, E. (2022). Global population and conservation status of the Great Black-backed Gull Larus marinus. Bird Conservation International p.1-11. DOI: https://doi.org/10.1017/S0959270922000181.
Ludwig, S.C., McCluskie, A., Keane, P., Barlow, C., Francksen, R.M., Bubb, D., Roos, S., Aebischer, N.J., & Baines, D. (2018). Diversionary feeding and nestling diet of Hen Harriers Circus cyaneus. Bird Study 65: 431-443.
Luxmoore, R., Swann, R., and Bell, E., (2019). Canna seabird recovery project: 10 years on. In: Veitch, C.R., Clout, M.N., Martin, A.R., Russell, J.C., & West, C.J. (eds) (2019). Island invasives; scaling up to meet the challenge, pp. 576-579. Occasional Paper SSC no. 62. Gland, Switzerland: IUCN.
MacArthur Green (2021). Report to Crown Estate Scotland and SOWEC: HRA Derogation Scope B – Review of seabird strategic compensation options. Crown Estate Scotland, Edinburgh: 166 pp.
MacDonald, A., Heath, M.R., Greenstreet, S.P.R. & Speirs, D. (2019). Timing of sandeel spawning and hatching off the east coast of Scotland. Frontiers in Marine Science 6, 70.
Mallord, J. W., Dolman, P. M., Brown, A. F. & Sutherland, W. J. (2007) Linking recreational disturbance to population size in a ground-nesting passerine. Journal of Applied Ecology, 44, 185-195.
Mason, L., Green, R.E., Hirons, G.J.M., Skinner, A.M.J., Peault, S.C., Upcott, E.V., Wells, E., Wilding, D.J., & Smart, J. (2021). Experimental diversionary feeding of red kites Milvus milvus reduces chick predation and enhances breeding productivity of northern lapwings Vanellus vanellus. Journal for Nature Conservation, 64, https://doi.org/10.1016/j.jnc.2021.126051.
Massaro, M., Chardine, J.W., & Jones, I.L. (2001), Relationships between Black-Legged Kittiwake Nest-Site Characteristics and Susceptibility to Predation by Large Gulls. The Condor: 103 (4): 793-801.
Mavor, R.A., Parsons, M., Heubeck, M. & Schmitt, S. (2004). Seabird numbers and breeding success in Britain and Ireland, 2003. UK Nature Conservation No. 28, Joint Nature Conservation Committee, Peterborough:100 pp.
Mcdonald, R., & Hutchings, M.R. (1997). The stats of ship rats Rattus rattus on the Shiant Islands, Outer Hebrides, Scotland. Biological Conservation 82(1):113-117.
Mitchell, I.P., Newton, S.F., Ratcliffe, N., & Dunn, T.E. (2004). Seabird populations of Britain and Ireland. A & C Black Publishers Ltd, London. ISBN: 0-7136-6901-2.
Mitchell, I., Daunt, F., Frederiksen, M., & Wade, K. (2020). Impacts of climate change on seabirds, relevant to the coastal and marine environment around the UK. MCCIP Science Review 2020, p.382-399.
Moors, P.J. (1985). Norway rats (Rattus norvegicus) on the Noises and Motukawo Islands, Hauraki Gulf, New Zealand. New Zealand Journal of Ecology 8: 37-54.
Morris, R. (2003). The Wildlife of Inchcolm. Hillside, Edinburgh. ISBN: 0-9544760-1-8.
Murray, D.S. (2015). The Guga Hunters. Birlinn, Edinburgh, UK. ISBN: 9780857907653. 256pp.
Murray, S., Harris, M.P. & Wanless, S. (2014). The Bass Rock – now the world’s largest Northern Gannet colony. British Birds 107 765-769.
Murison, G. (2002) The impact of human disturbance on the breeding success of nightjar Caprimulgus europaeus on heathlands in south Dorset, England. English Nature, Peterborough.
Nelson, R.W. (1990). Status of the Peregrine Falcon Falco peregrinus pealei, on Langara Island, Queen Charlotte Islands, British Columbia, 1968-1989. The Canadian Field-Naturalist, 104: 193-199.
Nelson, J.B. (2002). The Atlantic Gannet. Fenix Books Limited, Great Yarmouth.
Nelson, R.W. (2004). The 1995 Peregrine Falcon survey on Langara Island. The 1995 Peregrine Falcon survey in Canada. Number 110, Canadian Wildlife Service.
Nelson, R.W. & Myres, M.T. 1976. Declines in populations of peregrine falcons and their seabird prey at Langara Island, British Colombia. The Condor 78: 281-293.
Nettleship, D.N. (1972). Breeding success of the Common Puffin (Fractercula arctica L.) on different habitats at Great Island, Newfoundland. Ecological Monographs 42:239-268.
Northridge, S., Kingston, A. & Coram, A. (2020). Preliminary estimates of seabird bycatch by UK vessels in UK and adjacent waters. Final Report to JNCC (for Defra report ME6024 rev). Scottish Ocean Institute, University of St Andrews.
O’Hanlon, N., Bond, A.L., Masden, E., Lavers, J.L. & James, N.A. (2021). Measuring nest incorporation of anthropogenic debris by seabird: An opportunistic approach increases geographic scope and reduces costs. Marine Pollution Bulletin 171: https://doi.org/10.1016/j.marpolbul.2021.112706
Oliveira, N., Henriques, A., Miodonski, J., Pereira, J., Marujo, D., Almeida, A., Barros, N., Andrade, J., Marçalo, A., Santos, J., Oliveira, I.B., Ferreira, M., Araújo, H., Monteiro, S., Vingada, J. & Ramírez, I. (2015). Seabird bycatch in Portuguese mainland coastal fisheries: An assessment through on-board observations and fishermen interviews. Global Ecology and Conservation 3: 51–61.
Paine, R.T., Wootton, J.T. & Boersma, P.D. (1990). Direct and indirect effects of Peregrine Falcon predation on seabird abudance. The Auk 107: 1-9.
Palmer, A., Nordmeyer, D.L. & Roby D.D. (2004). Nestling provisioning rates of Peregrine Falcons in interior Alaska. Journal of Raptor Research 38(1): 9–18.
Parker, M.W., Kress, S.W., Golighty, R.T., Carter, H.R., Parsons, E.B., Schubel. S.E., Boyce, J.A., McChesney, G.J. and Wisely, S.M., (2007), Assessment of social attraction techniques used to restore a Common Murre colony in central California, Waterbirds: The International Journal of Waterbird Biology, 30:17-28.
Parrish, J,K., and Paine, R.T. (1996). Ecological interactions and habitat modification in nesting Common Murres Uria aalge. Bird Conservation International, 6:261-269.
Perkins, A., Ratcliffe, N, Suddaby, D., Ribbands, B, Smith, C., Ellis, P., Meek, E. & Bolton, M. (2018). Combined bottom-up and top-down pressure drive catastrophic population declines of Arctic skuas in Scotland. Journal of Animal Ecology, 87(6), 1753-1586.
Pierce, K.E., Harris, R.J., Larnad, L.A. & Pokas, M.A. (2004). Obstruction and starvation associated with plastic ingestion in a Northern Gannet Morus bassanus and a Greater Shearwater Puffinus gravis. Marine Ornithology 32: 187–189.
Pierotti, R. (1983). Gull-Puffin interactions on Great Island, Newfoundland. Biological Conservation 26:1-14.
Pierrotti, R., & Annett, C.A. (1991). Diet choice in the Herring Gull: constraints imposed by reproductive and ecological factors. Ecology, 71: Issue 1: 319-328.
Ratcliffe, N., Mitchell, I., Varnham, K., Verboven, N., & Higson, P. (2009). How to prioritise rat management for the benefit of petrels: a case study of the UK, Channel Islands and Isle of Man. Ibis 151: 699-708.
Redpath, S.M., Thirgood, S.J., & Leckie, F.M. (2001). Does supplementary feeding reduce predation of red grouse by hen harriers? Journal of Applied Ecology 38:1157-1168.
Redpath, S. & Thirgood, S. (2009). Hen harriers and red grouse: moving towards consensus? Journal of Applied Ecology 46, 961-963.
Regehr, H.M. & Montevecchi, W.A. (1996). Interactive effects of food shortage and predation on breeding failure of black-legged kittiwakes: effects of fisheries activities and implications for indicator species. Marine ecology progress series 155, 249-260.
Remacha, C., Delgado J.A., Bulaic M., Pérez-Tris J. (2016) Human Disturbance during Early Life Impairs Nestling Growth in Birds Inhabiting a Nature Recreation Area. Plos one, DOI: 10.1371/journal.pone.0166748
Roman, L., Kastury, F., Petit, S., Aleman, R., Wilcox, C., Hardesty, B.D., & Hindell, M.A. (2020). Plastic, nutrition and pollution; relationships between ingested plastic and metal concentrations in the livers of two Pachyptila seabirds. Nature Scientific Reports 10: 10823. https://doi.org/10.1038/s41598-020-75024-6.
Roos, S., Smart, J., Gibbons, D.W., & Wildon, J.D. (2018). A review of predation as a limiting factor for bird populations in mesopredator-rich landscapes: a case study of the UK. Biological Reviews doi: 10.1111/brv.12426
RSPB News from the Rock No. 82. Available online at: https://community.rspb.org.uk/placestovisit/ramseyisland/b/ramseyisland-blog/posts/news-from-the-rock-82---seabird-count-update-razorbills.
Ruhlen, T. D., Abbott, S., Stenzel, L. E. & Page, G. W. (2003) Evidence that human disturbance reduces Snowy Plover chick survival. Journal of Field Ornithology, 74, 300-304.
Russell, J.C., Beaven, B.M., MacKay, J.W.B, Towns, D.R. & Clout M.N (2008). Testing island biosecurity systems for invasive rats. Wildlife Research 35, 215-221.Russell, J.C., Clout, M.N & Towns, D.R. (2007). Early detection and eradication of invading rats. USDA National Wildlife Research Center Sympoia.
Sanz-Aguillar, A., Martinez-Abrain, A., Tavecchia, G., Minguez, E. & Oro, D. (2009). Evidence-based culling of a facultative predator: Efficiacy and efficiency components. Volume 142, Issue 2, p. 424-431.
Sawyer, S.L., (2013), Establishment of a new breeding colony of Australasian gannets (Morus serrator) at Young Nick’s Head Peninsula, Notornis, 60: 180-182
Scottish Wildlife Trust (1998). Handa Island Wildlife Reserve: Ranger’s Report 1998.
Scottish Wildlife Trust (1999). Handa Island Wildlife Reserve: Ranger’s Report 1999.
Scottish Wildlife Trust (2001). Handa Island Wildlife Reserve: Ranger’s Report 2001.
Scottish Wildlife Trust (2001). Handa Island Wildlife Reserve: Ranger’s Report 2002.
Scottish Wildlife Trust (2003). Handa Island Wildlife Reserve: Ranger’s Report 2003.
Scottish Wildlife Trust (2005). Handa Island Wildlife Reserve: Ranger’s Report 2005.
Scottish Wildlife Trust (2016). Handa Island Wildlife Reserve: Ranger’s Report 2016.
Scottish Wildlife Trust (2017). Handa Island Wildlife Reserve: Ranger’s Report 2017.
Scottish Wildlife Trust (2018). Handa Island Wildlife Reserve: Ranger’s Report 2018.
Scottish Wildlife Trust (2019). Handa Island Wildlife Reserve: Ranger’s Report 2019.
Scottish Wildlife Trust (2020). Handa Island Wildlife Reserve: Ranger’s Report 2020.
Scottish Wildlife Trust (2021). Handa Island Wildlife Reserve: Ranger’s Report 2021.
Searle, K.R., Regan, C.E., Perrow, M.R., Butler, A., Rindorf, A., Harris, M.P., Newell, M.A., Wanless, S., & Daunt, F. (in prep). Effects of a fishery closure and prey abundance on seabird diet and breeding success: implications for strategic fisheries management and seabird conservation. Submitted to Biological Conservation.
Smart, J. & Amar, A. (2018). Diversionary feeding as a means of reducing raptor predation at seabird breeding colonies. Journal for Nature Conservation 46: 48–55.
Stanbury, A., Thomas, S., Aergeter, J., Brown, A., Bullock, D., Eaton, M., Lock, L., Luxmoore, R., Roy, S., Whittaker, S., & Oppel, S. (2017). Prioritising islands in the UK and crown dependencies for the eradication of invasive alien vertebrates and rodent biosecurity. Eur J Wildl Res DOI 101.007s10344-017-1084-7.
Stillman, R. A., West, A. D., Caldow, R. W. G. & Durell, S. E. A. L. V. D. (2007) Predicting the effect of disturbance on coastal birds. Ibis, 149, 73-81.
Stoneman, J.G. & Willcox, N.A. (1995). Seabirds of Handa. Scottish Birds 18:87-87.
Stoneman, J.G. & Zonfrillo, B. (2005). The eradication of Brown Rats from Handa Island, Sutherland. Scottish Birds 25:17-23.
Storrier, K.L. (2004). The Forth Coastal Litter Campaign: Working Towards a Litter-free Forth. Forth Estuary Forum, Rosyth, Scotland: 72 pp.
Stroud, D.A., Bainbridge, I.P., Maddock, A., Anthony, S., Baker, H., Buxton, N., Chambers, D., Enlander, I., Hearn, R.D., Jennings, K.R, Mavor, R., Whitehead, S. & Wilson, J.D. - on behalf of the UK SPA & Ramsar Scientific Working Group (eds.) (2016). The status of UK SPAs in the 2000s: the Third Network Review. [c.1,108] pp. JNCC, Peterborough.
Sutton, L.J. & Loram, S.W. (2021). Diet specialisation in an insular population of coastal Peregrine Falcons. bioRev prepint doi: https://doi.org/10.1101/2021.09.07.459223.
Swan, G.J.F., Redpath, S.M., Bearhop, S., & McDonald, R.A. (2017). Ecology of problem individuals and the efficacy of selective wildlife management. Trends in Ecology and Evolution Vol. 32: 7, p.518-530.
STR. (2022). East Lothian Visitor Survey. Final Report by STR prepared for East Lothian Council. Accessed June 2022: East Lothian Visitor Survey 2022 | East Lothian Council.: https://www.eastlothian.gov.uk/downloads/file/28174/east_lothian_visitor_survey_2021
Tabak, M.A., Poncet, S., Passfield, K., Goheen, J.R. & Martinez del Rio, C. (2015). Rat eradication and the resistance and resilience of passerine bird assemblages in the Falkland Islands. Journal of Animal Ecology 84: 755-764.
The Forth Seabird Group (2019). The Forth Islands Bird Report 2018. Compiled by Bill Bruce.
The Seabird Group (2009). Newsletter 112. Ailsa Craig. Accessible online at: http://www.seabirdgroup.org.uk/newsletters/
The Seabird Group (2019). Newsletter 142. Canna. Accessible online at: http://www.seabirdgroup.org.uk/newsletters/
Thomas, S., & Varnham, S. (2016). Island Biosecurity Manual. Seabird Island Restoration Project, RSPB. RSPB, Sandy, Bedfordshire.
Thomas, S., Brown, A., Bullock, D., Lock, L. Luxmoore, R., Roy, S., Stanbury, A., & Varnham, K. (2017a). Island restoration in the UK – past, present and future. British Wildlife 28:4, p.231-243.
Thomas, S., Varnham, K., & Harvey, S. (2017b). Current Recommended Procedures for UK (bait station) rodent eradication project. (Version 4.0). RSPB, Sandy, Bedfordshire.
Travers, T., Lea, M.A., Alderman, R., Terauds, A. & Shaw, J. (2020). Bottom-up effect of eradications: The unintended consequences for top-order predators when eradicating invasive prey. Journal of Applied Ecology, 58:4 pp. 801-811.
Trinder, M. (2016). Population viability analysis of the Sula Sgeir gannet population. Scottish Natural Heritage Commissioned Report No. 897, Scottish Natural Heritage, Invervess: 21pp.
Tyson, C., Shamoun-Baranes, J., Van Loon, E., Camphuysen, K., & Hintzen, N. (2015). Individual specialisation on fishery discards by Lesser Black-backed Gulls (Larus fuscus). ICES Journal of Marine Science, Volume 72, Issue 6, p. 1882-1891.
Veitch, B.G., Robertson, G.J., Jones, I.L & Bond, A.L. (2016). Great Black-backed Gull (Larus marinus) predation on seabird populations at two colonies in Eastern Canada. Waterbirds 39: 235-245.
Verhoeven, Mo A., Jelle Loonstra, A.H., Pingle, T., Kasperma, W., Whiffin, M., McBride, A., Sjoerdsma, P., Roodhard, C., Burgess, M.D, Piersma, T., Smart, Jennifer. (2022). Do ditch-side electric fences improve the breeding productivity of ground-nesting waders? Ecological Solutions and Evidence, 3 (2). 13pp. https://doi.org/10.1002/2688-8319.12143.
Votier, S.C., Furness, R.W., Bearhop, S., Crane, J.E., Caldow, R.W., Catry, P., Ensor, K., Hamer, K.C., Hudson, A.V., Kalmbach, E., Klomp, N.I., Pfeiffer, S., Phillips, R.A., Prieto, I., & Thompson, D. (2004a). Changes in fisheries discard rates and seabird communities. Nature, Vol. 427, p. 727-730.
Votier, S.C., Bearhop, S., Ratcliffe, N., Phillips, R.A., & Furness, R.W. (2004b). Predation by great skuas at a large Shetland seabird colony. Journal of Applied Ecology 41, 1117-1128.
Votier, S.C., Archibald, K., Morgan, G. & Morgan, L. (2011). The use of plastic debris as nesting material by a colonial seabird and associated entanglement mortality. Marine Pollution Bulletin 62: 168–172.
Wanless, S. (1983). Seasonal variation in the numbers and condition of Gannets Sula bassana dying on Ailsa Craig, Scotland. Bird Study 30(2): 102–108.
Wanless, S., Murray, S., & Harris, M.P. (2005). The Status of Northern Gannet in Britain & Ireland in 2003/4. British Birds 98: 280-294.
Wanless, S., Harris, M.P., Newell, M.A., Speakman, J.R. & Daunt, F. (2018). Community-wide decline in the occurrence of lesser sandeels Ammodytes marinus in seabird chick diets at a North Sea colony. Marine Ecology Progress Series 600: 193–206.
Watson, H., Bolton, M., Monaghan, P. (2014). Out of sight but not out of harm’s way: Human disturbance reduces reproductive success of a cavity-nesting seabird. Biological Conservation 174:127-133.
Wernham, C.V., Toms, M.P, Marchant, J.H., Clark, J.A., Sririwardena, G.M., Baillie, S.R. (eds) (2002). The Migration Atlas: movements of the birds of Britain and Ireland. T. & A.D. Poyser, ISBN: 978-0713665147.
West, A. D., Goss-Custard, J. D., Stillman, R. A., Caldow, R. W. G., Durell, S. & McGrorty, S. (2002) Predicting the impacts of disturbance on shorebird mortality using a behaviour-based model. Biological Conservation, 106, 319-328.
Westerberg, K., Brown, R., Eagle, G & Votier, S.C. (2019). Intra-population variation in the diet of an avian top predator: generalist and specialist foraging in Great Black-backed Gulls Larus marinus. Bird Study, https://doi.org/10.1080/00063657.2019.1693961.
White, J., Leclaire, S., Kriloff, M., Mulard, H., Hatch, S.A. & Danchin, E. (2010). Sustained increase in food supplies reduces broodmate aggression in black-legged kittiwakes. Animal Behaviour 79:1095-1100.
Wilson, M (2018). Pylons and power stations. BTO News, Spring 2018, p.20-21.
Witmer, G.W., Nolte, D.L., Stewart, W.B. (2000). Integrated pest management of black bear reforestation damage. In: Salmon, T.P, Crabb, A.C. (eds). Proceedings of the Nineteenth Vertebrate Pest Conference. San Diego, California, 6-9 March 2000. University of California, pp. 228-235.
Yasue, M. & Dearden, P. (2006) The potential impact of tourism development on habitat availability and productivity of Malaysian plovers Charadrius peronii. Journal of Applied Ecology, 43, 978-989.
Ziegltrum, G.J. (2008). Impacts of the black bear supplemental feeding program on ecology in western Washington. Human-Wildlife Conflicts 2:153-159.
APPENDIX 1
Table A2. Islands on the east coast of the UK from RSPB Islands Database that also support breeding colonies of the key species (identified from JNCC Seabird Nesting Counts). Highlighted sites show islands that support at least some of the key species and have no biosecurity measures in place.
FID | X | Y | Site name/description | Kittiwake AON | Guillemot (inds) | Razorbill (inds) | Puffin AOB | Gannet AON | Rodent status | Other information |
|---|---|---|---|---|---|---|---|---|---|---|
1417 | 4419014 | 564949 | Marsden Rock | 2472 | 0 | small colony | 0 | 0 | May not be rats - cliff faces extremely steep | Joined to mainland at LW. |
1480 | 429365 | 604643 | Coquet Island | 51 | 0 | 0 | 17208 | 0 | No rats, biosecurity plan in place | Not suitable |
1501 | 424706 | 638991 | Longstone (Farnes) |
|
| 2 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1510 | 423769 | 637722 | Brownsman and Staple (Farnes) | 2406 | 20015 | 67 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1511 | 424023 | 638540 | Harcar (Farnes) | 113 | 117 | 8 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1514 | 423758 | 638170 | Roddam and Green (Farnes) | 30 | 170 | 0 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1522 | 423468 | 638393 | Wamses (Farnes) | 157 | 1483 | 15 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1523 | 422818 | 636015 | Skeney Scar (Farnes) | 121 | 1306 | 12 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1528 | 422141 | 636128 | Inner Farne, Wideopens, Knoxes | 2246 | 8196 | 103 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
1531 | 420564 | 637302 | Megstone (Farnes) | 23 | 210 | 2 | 0 | 0 | No rats, biosecurity plan in place | Not suitable |
2001 | 368152 | 679585 | Dunbar – includes rocky outcrops offshore | 808 | 0 | 0 | 0 | 0 | Most of colony on mainland, a few Kittiwakes nesting on outcrops. | See section 4 |
2035 | 365559 | 699428 | Isle of May (Forth Islands) | 5193 | 18,156 | 4813 | 39200 | 0 | No rats, biosecurity plan in place | Not suitable |
2121 | 360225 | 687391 | Bass Rock (Forth Islands) | 1307 | 2422 | 116 | 0 | 44110 | No rats, biosecurity plan in place | Not suitable |
2156 | 355281 | 686995 | Craigleith (Forth Islands) | 511 | 1708 | 186 | 28000 |
| No rats, biosecurity plan in place | Not suitable |
2158 | 353482 | 686596 | Lamb (Forth Islands) | 115 | 3760 | 101 | 7 |
| No rats, biosecurity plan in place | Not suitable |
2172 | 351283 | 686841 | Fidra (Forth Islands) | 225 | 376 | 161 | 405 |
| No rats, biosecurity plan in place | Not suitable |
2278 | 329391 | 682653 | Inchkeith | 349 | 48 | 85 | 1641 |
| Some evidence of rats | Suitable |
2343 | 320706 | 680438 | Inchmickery | 0 | 0 | 0 | 0 | 22 | No rats, biosecurity plan in place | Not suitable |
2385 | 318935 | 682544 | Inchcolm | 116 | 0 | 5 | 40 | 0 | Black rat present | Suitable |
Table A3. Islands in the UK supporting gannetries and breeding colonies of the other key species.
Site | Subsite | Gannet AON | Kittiwake AON | Guillemot (inds) | Razorbill (inds) | Puffin AOB | Suitability for rat removal |
|---|---|---|---|---|---|---|---|
BULL ROCK* | THE BULL | 1879 | 0 | 0 | 0 | 0 | SPA, no evidence of rats |
IRELAND'S EYE* | IRELAND'S EYE 2 | 147 | 809 | 1336 | 202 | 0 | SPA, no evidence of rats |
LITTLE SKELLIG* | LITTLE SKELLIG - WHOLE ISLAND | 28799 | 250 | 1129 | 68 | 0 | SPA, IBA, no evidence of rats |
CLARE ISLAND* | CLARE ISLAND 12 | 3 | 503 | 970 | 190 | 25 | SPA, no evidence of rats |
GREAT SALTEE* | GREAT SALTEE ISLAND | 1930 | 2125 | 21436 | 3239 | 1522 | SPA, rat removal plan underway |
CHANNEL ISLANDS | LES ETACS | 3450 | 0 | 0 | 0 | 0 | No rats |
CHANNEL ISLANDS | ORTAC | 2500 | 0 | 0 | 0 | 0 | No rats |
FORTH ISLANDS - BASS ROCK TO HAYSTACK | BASS ROCK | 44110 | 1307 | 2422 | 116 | 0 | SPA, biosecurity measures already underway |
AILSA CRAIG | AILSA CRAIG (WHOLE ISLAND) | 35825 | 1675 | 9415 | 1471 | 20 | SPA, rats already removed |
SULE STACK | SULE STACK1 | 5137 | 51 | 1062 | 10 | 0 | SPA, no rats |
FAIR ISLE | WHOLE ISLAND | 1123 | 8204 | 39257 | 3599 | 40000 | No rats |
FOULA | FOULA 8 | 603 | 1061 | 22874 | 369 | 0 | SPA, biosecurity measures already underway |
FOULA | FOULA 9 | 120 | 346 | 4730 | 116 | 0 | SPA, biosecurity measures already underway |
NOSS NNR | NOSS | 8017 | 2395 | 45777 | 1984 | 1892 | SPA, biosecurity measures already underway |
BORERAY, ST KILDA | BORERAY GANNETS | 61340 | 0 | 0 | 0 | 0 | SPA, rats already removed |
FLANNAN ISLES | ROAREIM | 1244 | 0 | 0 | 0 | 0 | SPA, biosecurity measures already underway |
SULA SGEIR ISLAND | SULA SGEIR | 10703 | 1206 | 20877 | 801 | 177 | SPA, no rats present |
MONRIETH CLIFFS & SCAR ROCKS | BIG SCAR | 1670 | 4 | 1753 | 39 | 0 | RSPB reserve, no rats |
GRASSHOLM, BISHOP & CLERKS, AND RAMSEY | GRASSHOLM | 30688 | 7 | 1346 | 31 | 0 | SPA, biosecurity measures already underway |
*SPAs in the Republic of Ireland are not covered under the Biosecurity LIFE Project.
Table A4. Top Twenty islands in the UK for Guillemot based on abundance.
Rank | Site | Subsite | Guillemot | Razorbill | Puffin | Kittiwake |
|---|---|---|---|---|---|---|
1 | HANDA ISLAND | WHOLE ISLAND | 112676 | 16991 | 735 | 7013 |
2 | RATHLIN ISLAND | WHOLE ISLAND | 95117 | 20860 | 1579 | 9917 |
3 | LAMBAY ISLAND | WHOLE ISLAND | 60754 | 4337 | 289 | 4091 |
4 | NOSS | WHOLE ISLAND | 45777 | 1984 | 1892 | 2395 |
5 | FOULA | WHOLE ISLAND | 41435 | 2121 | 22500 | 1934 |
6 | FAIR ISLE | WHOLE ISLAND | 39257 | 3599 | 40000 | 8204 |
7 | FARNE ISLANDS | ALL ISLANDS | 31497 | 209 | 55674 | 5096 |
8 | ISLE OF MAY | WHOLE ISLAND COUNT | 28103 | 4114 | 42000 | 3639 |
9 | COLONSAY | WHOLE ISLAND | 26429 | 2742 | 1 | 6485 |
10 | ISLE OF COLONSAY | WHOLE ISLAND | 26429 | 2742 | 1 | 6485 |
11 | GREAT SALTEE | WHOLE ISLAND | 21436 | 3239 | 1522 | 2125 |
12 | SULA SGEIR | WHOLE ISLAND | 20877 | 801 | 177 | 1206 |
13 | COPINSAY | WHOLE ISLAND | 20045 | 671 | 350 | 4364 |
14 | BERNERAY | WHOLE ISLAND | 19083 | 16513 | 1979 | 2613 |
15 | SHIANT ISLANDS | ALL ISLANDS | 16456 | 8046 | 65170 | 2006 |
16 | DUNCANSBY HEAD TO SMOO (INCLUDES STROMA) - HIGHLAND | STROMA (WHOLE ISLAND) | 14760 | 280 | 26 | 821 |
17 | FLANNAN ISLES | ALL ISLANDS | 14638 | 1569 | 15761 | 1244 |
18 | SKOMER AND MIDDLEHOLM ISLANDS | SKOMER ISLAND | 13852 | 3898 | 7076 | 2257 |
19 | MINGULAY | WHOLE ISLAND | 13387 | 6387 | 3827 | 2898 |
20 | NORTH RONA | WHOLE ISLAND | 10497 | 824 | 5265 | 3398 |
Table A5. Top twenty islands for Kittiwake based on abundance.
Rank | Site | Subsite | Kittiwake | Guillemot | Razorbill | Puffin |
|---|---|---|---|---|---|---|
1 | BORERAY, ST KILDA | WHOLE ISLAND | 61340 | 5880 | 256 | 50999 |
2 | RATHLIN ISLAND | WHOLE ISLAND | 9917 | 95117 | 20860 | 1579 |
3 | FAIR ISLE | WHOLE ISLAND | 8204 | 39257 | 3599 | 40000 |
4 | HANDA ISLAND | WHOLE ISLAND | 7013 | 112676 | 16991 | 735 |
5 | COLONSAY | WHOLE ISLAND | 6485 | 26429 | 2742 | 1 |
6 | ISLE OF COLONSAY | WHOLE ISLAND | 6485 | 26429 | 2742 | 1 |
7 | FARNE ISLANDS | ALL ISLANDS | 5096 | 31497 | 209 | 55674 |
8 | COPINSAY | WHOLE ISLAND | 4364 | 20045 | 671 | 350 |
9 | LAMBAY ISLAND | WHOLE ISLAND | 4091 | 60754 | 4337 | 289 |
10 | ISLE OF MAY | WHOLE ISLAND COUNT | 3639 | 28103 | 4114 | 42000 |
11 | NORTH RONA | WHOLE ISLAND | 3398 | 10497 | 824 | 5265 |
12 | MINGULAY | WHOLE ISLAND | 2898 | 13387 | 6387 | 3827 |
13 | ROUSAY | WHOLE ISLAND | 2713 | 6205 | 510 | 53 |
14 | BERNERAY | WHOLE ISLAND | 2613 | 19083 | 16513 | 1979 |
15 | NOSS | WHOLE ISLAND | 2395 | 45777 | 1984 | 1892 |
16 | SKOMER AND MIDDLEHOLM ISLANDS | SKOMER ISLAND | 2257 | 13852 | 3898 | 7076 |
17 | GREAT SALTEE | WHOLE ISLAND | 2125 | 21436 | 3239 | 1522 |
18 | SHIANT ISLANDS | ALL ISLANDS | 2006 | 16456 | 8046 | 65170 |
19 | FOULA | WHOLE ISLAND | 1934 | 41435 | 2121 | 22500 |
20 | AILSA CRAIG | WHOLE ISLAND | 1675 | 9415 | 1,471 | 20 |
Table A6. Top twenty islands for Razorbill based on abundance.
Rank | Site | Subsite | Razorbill | Kittiwake | Guillemot | Puffin |
|---|---|---|---|---|---|---|
1 | RATHLIN ISLAND | WHOLE ISLAND | 20860 | 9917 | 95117 | 1579 |
2 | HANDA ISLAND | WHOLE ISLAND | 16991 | 7013 | 112676 | 735 |
3 | BERNERAY | WHOLE ISLAND | 16513 | 2613 | 19083 | 1979 |
4 | SHIANT ISLANDS | ALL ISLANDS | 8046 | 2006 | 16456 | 65170 |
5 | MINGULAY | WHOLE ISLAND | 6387 | 2898 | 13387 | 3827 |
6 | LAMBAY ISLAND | WHOLE ISLAND | 4337 | 4091 | 60754 | 289 |
7 | ISLE OF MAY | WHOLE ISLAND COUNT | 4114 | 3639 | 28103 | 42000 |
8 | SKOMER AND MIDDLEHOLM ISLANDS | SKOMER ISLAND | 3898 | 2257 | 13852 | 7076 |
9 | FAIR ISLE | WHOLE ISLAND | 3599 | 8204 | 39257 | 40000 |
10 | GREAT SALTEE | WHOLE ISLAND | 3239 | 2125 | 21436 | 1522 |
11 | COLONSAY | WHOLE ISLAND | 2742 | 6485 | 26429 | 1 |
12 | ISLE OF COLONSAY | WHOLE ISLAND | 2742 | 6485 | 26429 | 1 |
13 | FOULA | WHOLE ISLAND | 2121 | 1934 | 41435 | 22500 |
14 | NOSS | WHOLE ISLAND | 1984 | 2395 | 45777 | 1892 |
15 | FLANNAN ISLES | ALL ISLANDS | 1569 | 1244 | 14638 | 15761 |
16 | SANDA ISLAND, SHEEP ISLAND AND GLUNIMORE ISLAND | SHEEP ISLAND | 1500 | 0 | 40 | 78 |
17 | GRASSHOLM, BISHOP & CLERKS, AND RAMSEY | RAMSEY ISLAND | 1499 | 293 | 3284 | 0 |
18 | AILSA CRAIG | WHOLE ISLAND | 1471 | 1675 | 9415 | 20 |
19 | SKOKHOLM | WHOLE ISLAND | 1234 | 0 | 996 | 2055 |
20 | TORY ISLAND AND BLOODY FORELAND | TORY ISLAND | 1002 | 408 | 568 | 1402 |
Table A7. Top twenty islands for Puffin based on abundance.
Rank | Site | Subsite | Puffin | Guillemot | Razorbill | Kittiwake |
|---|---|---|---|---|---|---|
1 | SHIANT ISLANDS | ALL ISLANDS | 65170 | 16456 | 8046 | 2006 |
2 | SULE SKERRY | WHOLE ISLAND | 59471 | 10331 | 88 | 1275 |
3 | FARNE ISLANDS | ALL ISLANDS | 55674 | 31497 | 209 | 5096 |
4 | BORERAY, ST KILDA | WHOLE ISLAND | 50999 | 5880 | 256 | 61340 |
5 | ISLE OF MAY | WHOLE ISLAND COUNT | 42000 | 28103 | 4114 | 3639 |
6 | FAIR ISLE | WHOLE ISLAND | 40000 | 39257 | 3599 | 8204 |
7 | FORTH ISLANDS - BASS ROCK TO HAYSTACK | CRAIGLEITH | 28000 | 1708 | 186 | 511 |
8 | SOAY, ST KILDA | WHOLE ISLAND | 27600 | 4404 | 197 | 0 |
9 | FOULA | WHOLE ISLAND | 22500 | 41435 | 2121 | 1934 |
10 | COQUET ISLAND | WHOLE ISLAND | 17208 | 0 | 0 | 0 |
11 | FLANNAN ISLES | ALL ISLANDS | 15761 | 14638 | 1569 | 1244 |
12 | SKOMER AND MIDDLEHOLM ISLANDS | SKOMER ISLAND | 7076 | 13852 | 3898 | 2257 |
13 | NORTH RONA | WHOLE ISLAND | 5265 | 10497 | 824 | 3398 |
14 | PUFFIN ISLAND - KERRY | WHOLE ISLAND | 5125 | 92 | 35 | 0 |
15 | GREAT SKELLIG | WHOLE ISLAND | 4000 | 1422 | 386 | 0 |
16 | MINGULAY | WHOLE ISLAND | 3827 | 13387 | 6387 | 2898 |
17 | SKOKHOLM | WHOLE ISLAND | 2055 | 996 | 1234 | 0 |
18 | BERNERAY | WHOLE ISLAND | 1979 | 19083 | 16513 | 2613 |
19 | NOSS | WHOLE ISLAND | 1892 | 45777 | 1984 | 2395 |
20 | TRESHNISH ISLES | LUNGA (WHOLE ISLAND) | 1738 | 9433 | 943 | 782 |
.

Figure A9: Map of Handa showing Gaelic place names
Please note no mapping was carried out in 2019.
[1] Meeting held 26th November 2021.
[2] Email from NatureScot sent to SSER dated 26th April 2022.
[3] Https://opendata-rspbn.opendata.arcgis.com
[4] Seabird Nesting Counts (British Isles) - data.gov.uk
[5] SWT 2021. Handa Island Wildlife Reserve. Ranger’s Report 2021.
[6] Forth Islands Heritage Group 2021.
[7] Defined as being in the top twenty sites for that species, based on size of breeding colony.
[8] RSPB Priority list taken from Table 4 in LIFE13 NAT/UK/00209. Final Report. Covering the project activities from 01/10/2013 to 31/12/2018. Protecting and restoring the Shiant Isles SPA through rat removal, and safeguarding other seabird island SPAs in the UK.
[9] The target species for Inchcolm are Kittiwake, Razorbill, Puffin and Guillemot. Whilst Guillemot do not currently nest on Inchcolm the habitat is considered suitable.
[10] This includes the 10 birds observed in the 2021 survey conducted by the Forth Islands Heritage Group (FIHG 2021).
[11] Celebrating Seabird Success on the Island of Lundy and the Isles of Scilly - RSPB England - Our work - The RSPB Community
[12] Ailsa Craig - A Puffin Success Story - Argyll Cruising
[13] https://webgate.ec.europa.eu/life/publicWebsite/index.cfm?fuseaction=search.dspPage&n_proj_id=5018
[14] https://webgate.ec.europa.eu/life/publicWebsite/index.cfm?fuseaction=search.dspPage&n_proj_id=5018
[15] Celebrating Seabird Success on the Island of Lundy and the Isles of Scilly - RSPB England - Our work - The RSPB Community
[16] It is possible that the arrival of hedgehogs in 2002 and their subsequent increase may have been responsible, although this cannot be proven.
[17] Two-tailed test, n=6, Test statistic =0, Critical value= 0, p=0.05.
[18] Two-tailed test, n=6, Test statistic =0, Critical value= 0, p=0.05.
[19] These values vary very little over time.
[20] https://www.ayrshire-birding.org.uk/2001/01/ailsa_craig_before_and_after_the_eradication_of_rats_in_1991/
[21] RSPB Scotland Comments on Berwick Bank Wind Farm: Meeting 4, 21st September 2022.
[23] Email from NatureScot sent to SSER dated 26th April 2022.
[24] NatureScot advice on questions from 8th June derogation case meeting, letter received 6th July 2022.
[25] Marine Scotland Science responses to questions issued by Berwick Bank at the 3rd Compensatory measures meeting (held 8 June 2022) (Letter dated 12 July 2022).
[26] https://jncc.gov.uk/our-work/black-legged-kittiwake-rissa-tridactyla/
[27] It should be noted in 1987 individual sections were not counted but an overall total count was obtained from the East Lothian Bird Report
[28] Email from NatureScot sent to SSER dated 26th April 2022.
[29] It should be noted that the nests on the Sea wall (10) are regularly flooded out by storms. The numbers refer back to the locations marked in green on Figure 4.2
[30] This initiative involves giving fishing boats big bags to collect the plastics, ghost gear and other debris that gathers in their nets during normal fishing activities. When the fishing boats come into port, they unload the bags of litter. Rubbish is recycled or disposed of on land.
[31] Marine Scotland Science responses to questions issued by Berwick Bank at the 3rd Compensatory measures meeting (held 8 June 2022)
[32] The only exception to this being the Gannet harvest at Sula Sgeir.
[33] Although anecdotal evidence suggests that Gannet nests also suffer predation from Great-Black Backed Gull, the density and sheer number of Gannet nests on Bass Rock means that working on this species at this site would be prohibitively difficult.
[34] Great Black-backed Gull may soon be moved to the Red list due to severe population declines.
[35] Email from NatureScot sent to SSER dated 26th April 2022.
[36] An MSc student on Craigleith found a Puffin midden where a pair had stockpiled 30 corpses (The Forth Seabird Group 2019). Finney 2002 states that 3 specialist pairs on Isle of May took half the 191 Puffins found during corpse searches, giving an average of ~30 corpses per pair.
[37] Great Black-backed Gulls stash dead Puffins, often leaving the carcasses in pools to soften them.
[38] This is not enough Puffins to result in a population decline, to do this several thousand Puffins would need to be killed per year. (S. Lopez pers. comm.)
[39] In Great Skuas seabird predation increases in response to decreasing prey availability offshore (Votier et al 2004)
[40] Stealing fish from birds returning to the colony to feed chicks.
[41] Most recent counts presented in JNCC 2021.
[42] https://community.rspb.org.uk/placestovisit/minsmere/b/minsmere-blog/posts/life-and-death-at-minsmere
[43] The chick that is born second.
[44] https://www.nurdlehunt.org.uk/nurdle-finds.html
[45] NatureScot advice on questions from 8th June derogation case meeting, letter received 6th July 2022.