

1. Background
- Berwick Bank Wind Farm Limited (BBWFL) is a wholly owned subsidiary of SSE Renewables Limited and will hereafter be referred to as ‘the Applicant’. The Applicant is developing the Berwick Bank Wind Farm (hereafter referred to as ‘the Project’) located in the outer Forth and Tay region ( Figure 1.1 Open ▸ ).
- The Project is located adjacent to the consented Forth and Tay offshore wind farms consisting of Seagreen 1 and Seagreen 1A Project to the north, Inch Cape to the northwest and Neart na Gaoithe to the west. ( Figure 1.1 Open ▸ ).
- The marine components of the Project, which are the focus of this technical report, include the Proposed Development array area and the Proposed Development export cable corridor ( Figure 1.1 Open ▸ ), hereafter referred to collectively as the ‘Proposed Development’.
- The Project will, if consented, provide an estimated 4.1 GW of renewable energy, making it one of the largest offshore wind farms in the world. Given the anticipated operational life span of 35 years, the development will make a critical contribution to Scotland’s renewable energy target of 11 GW of new offshore wind by 2030. Initially, pre-July 2021, the area was named as two separate sites, Marr Bank and Berwick Bank, but these have now been merged to a single site, Berwick Bank Wind Farm.
- Wind turbine capacity is yet to be confirmed but is expected to be between 14 – 24 MW. 1. The Proposed Development will comprise up to 307 wind turbines, with the final number of wind turbines dependent on the capacity of individual wind turbines used, and also environmental and engineering survey results. The PDE considers a range of wind turbines with parameters reflective of potential generating capacities, allowing for a degree of flexibility to account for any anticipated developments in wind turbine technology while still allowing the production of the MDS for the assessment of effects. Importantly, the minimum lower blade tip height is 37 m (above LAT) for all wind turbine options as an engineering design measure to reduce collision risk to seabirds.
- The site boundary of the Proposed Development balances maximising the potential for renewable energy generation whilst reducing environmental impacts.
- The purpose of this technical report is to put the potential impacts from the Project on seabirds in the context of the wider ecosystem. This is in response to the earlier Scoping Opinion received in March 2021:
“there is a need to understand potential impacts holistically at a wider ecosystem scale rather than via the standard set of discrete individual receptor assessments. This assessment should focus on potential impacts across key trophic levels particularly in relation to the availability of prey species. This will enable a better understanding of the consequences (positive or negative) of any potential changes in prey distribution and abundance from the development of the wind farm on seabird and marine mammal (and other top predator) interests and what influence this may have on population level impacts.”
- NatureScot’s Scoping advice (December 2021) recommended that “further discussion is undertaken through the road map process to agree a suitable approach, including consideration of the upcoming OWEC PrePared project that is due to commence in 2022 and completed in 5 years”. The Ecosystem approach was discussed at the Berwick Bank Wind Farm Ornithology Road Map Meeting 4, on 31 January 2022, “The assessment should consider potential attraction to the turbine bases and how supporting processes are likely to change (e.g. physical processes). All of these dynamics should be considered through a climate change filter”.
Figure 1.1: Site boundaries for all consented and proposed wind farms currently within the Outer Firth of Forth.
- This report aims to review current literature and research on the impacts of climate change, commercial fisheries, offshore wind development, avian flu, invasive non-native species (INNS) and marine litter, which have been regarded as important threats to seabirds (Dias et al., 2019). More specifically, this report aims to:
- summarise the status of seabirds in the Forth of Tay region, and more widely across Scotland and the North Sea;
- describe the food web and associated trophic levels in the Proposed Development area;
- provide a literature review of likely significant effects on seabirds at an ecosystem-wide scale, including direct and indirect effects from climate change, commercial fisheries and offshore wind energy;
- provide a literature review of how climate change may impact seabirds in the future; and
- discuss the potential role of offshore wind energy in tackling climate and biodiversity emergencies.
- Following advice in the Scoping Opinion from MS-LOT (4 February 2022), this approach was agreed with NatureScot, RSPB Scotland and MSS through the Ornithology Road Map process (Road Map 5; Technical Appendix 11.8). This appendix reviews and presents available evidence primarily for the following seabird species, as they are relatively abundant in area of the Proposed Development and are sensitive to potential impacts from the Proposed Development as determined in volume 2 chapter 11:
- northern gannet (Morus bassanus, hereafter ‘gannet);
- common guillemot (Uria aalge, hereafter ‘guillemot’);
- herring gull (Larus argentatus);
- black-legged kittiwake (Rissa tridactyla, hereafter ‘kittiwake’);
- lesser black-backed gull (Larus fuscus);
- Atlantic puffin (Fratercula arctica, hereafter ‘puffin’); and
- razorbill (Alca torda).
2. Introduction
- Climate change is leading to environmental changes at all scales through rising global sea level, increasing frequency of severe weather events, and warming oceans (Walther, 2010). This is affecting the abundance and diversity of communities at all trophic levels, with potentially catastrophic consequences for some species and communities, including seabirds. A recent global assessment of threats to seabirds concluded that invasive non-native species (INNS), fisheries bycatch and climate change are the top three threats affecting seabirds globally (Dias et al. 2019). Energy production and mining, which included offshore wind was ranked ninth of 18 threats.
- In the UK, climate change is attributed to a significant decline in breeding abundance of seabirds by 20-30% since the early 1990s (Mitchell et al. 2018) which have primarily been attributed to direct (mortality due to extreme weather) and indirect (changes in prey availability) impacts (Mitchell et al. 2020).
- Rising energy demand from global markets in combination with the climate crisis has led to the rapid growth of the renewable energy sector, with renewable energy contributing to 22% of all energy consumed in Europe in 2020 (Eurostat, 2022). Offshore wind farms are an effective way to produce renewable energy, with the EU Offshore Renewable Energy Strategy setting targets of an installed capacity of 60GW from offshore wind in Europe by 2030 (European Commission, 2020). The UK has some of the best wind resources in Europe, contributing to it becoming one of the fastest growing renewable energy industries in the UK (DECC, 2011).
- Wind energy is a key renewable energy resource for Scotland, and it is a priority in the ‘Securing a green recovery on a path to net zero: climate change plan 2018–2032 – update’ to achieve delivery of greenhouse gas (GHG) emission reduction targets (Scottish Government, 2020a; Scottish Government, 2021). Carbon savings from offshore wind electricity generation compared to the same electricity produced from natural gas is on average 143 ktCO2e per wind turbine, which is equivalent to the CO2e emitted from approximately 70,000 UK petrol cars annually (ORE Catapult, 2021).
- Scotland has ambitious targets to reduce GHG emissions; the current Scottish climate change plan 2018-2032 (Scottish Government, 2020a) sets targets to reduce emissions by 75% by 2030 (compared with 1990) and to reach net zero by 2045, which has been written into law in the Climate Change (Scotland) Act 2009 (as amended by the Climate Change (Emissions Reduction Targets) (Scotland) Act 2019). These targets are also in line with Scotland’s commitments under the 2015 Paris Agreement to limit global average temperature increases to 1.5 degrees Celsius or less. The Scottish energy strategy sets a target for the equivalent of 50% of the energy for Scotland’s heat, transport and electricity consumption to be supplied by renewable sources by 2030 (Scottish Government, 2017; Scottish Government, 2021). Scotland’s National Marine Plan calls for the sustainable development of offshore wind and other renewable energy sources in most suitable locations, for both economic benefit and to achieve decarbonisation targets of 50g CO2/kWh by 2030 (Scottish Government, 2015).
- The Scottish Government’s sectoral marine plan for offshore wind energy supports the development of between 8GW and 11GW of offshore wind capacity by 2030 (Scottish Government, 2020b). The Scottish Government's energy and offshore wind policies are further discussed in volume 1, chapter 2.
- Offshore wind farms can affect seabirds through direct impacts, such as collisions, displacement, and barrier effects, as well as indirectly through impacts on the availability of their prey. These add to existing pressures from climate change and other threats. However, there is a balance to be struck between the potential impacts from the increasing number of offshore wind farms and reduced climate change impacts from a reduction in carbon emissions.
3. Status of seabirds
- Seabirds are mobile and generally wide-ranging species, able to exploit a range of environments from tropical to polar latitudes (Harrison et al., 2018; Dias et al., 2019). They play a key role in marine ecosystems and can indicate the health of an ecosystem. Globally, the status of seabirds is unfavourable, with the International Union for Convention of Nature (IUCN) ranking many species as critically endangered, endangered or vulnerable (BirdLife International, 2018), with approximately 50% of seabirds estimated as declining worldwide (Dias et al., 2019). Determining the status of global seabirds is challenging since they are generally highly mobile and transient (Dunn et al., 2019).
- Currently, 25 species of seabird breed around the UK (Mitchell et al., 2004). The Seabird 2000 census estimated over 8 million breeding seabirds were present around Britain and Ireland per year, although evidence of widespread declines in productivity (number of chicks fledged per pair) have since emerged which may be driving declines in breeding population size (JNCC 2022a). Data from the most recent census, Seabirds Count, should be available in 2023 (JNCC 2022b).
- Scotland hosts internationally important populations of seabirds (Mitchell et al., 2004), supporting an estimated 45% of Europe’s breeding seabirds (Forrester et al., 2007). Many species visit seasonally, migrating to exploit temporarily available food resources and advantageous weather conditions (Forrester et al., 2007). Approximately 58% and 46% of the northeast Atlantic and world gannet populations, respectively, are present in Scotland (Murray, Harris and Wanless, 2015) and 11% and 43% of Europe’s kittiwake and guillemot populations respectively are estimated to be present around the Scottish coast (Mitchell et al., 2004; Marine Scotland, 2020; JNCC, 2021a). Population estimates for puffin from the last UK seabird census estimated Scottish populations make up approximately 40% of the GB population (Mitchell et al., 2004; JNCC, 2021a).
- There has been an estimated 70% decline in worldwide monitored seabird populations between 1950 and 2010, which can likely be attributed to human interference and industrialisation (Paleczyny et al., 2015). The UK Marine Strategy Assessment (2019) described UK breeding seabirds as having ‘not achieved Good Environmental Status’, with several species reported as undergoing frequent and widespread breeding failure. Rates of decline were highest in the early 2010s, attributed to limited food availability and the presence of INNS species (Marine Scotland, 2020). Population declines have been observed in breeding seabirds in Scotland between 1986 – 2018 ( Figure 3.1 Open ▸ ).
- In the Marine Assessment of the Forth and Tay region, abundance and breeding success have shown a degree of stability over the period 2011 – 2018, although there were some concerns around kittiwake and shag populations (Marine Scotland, 2020; Figure 3.2 Open ▸ ).
- The abundance of gannets is increasing, including at Bass Rock, the closest colony to the Proposed Development, where the population is estimated to have increased by 53% between the census in 2003-04 and 2014 (from 49,098 to 75,259 individuals) (JNCC, 2021b). During baseline surveys, gannets were abundant within the Proposed Development area, especially during the breeding season (See Appendix 11.1: Baseline Ornithology Technical Report) and have been commonly observed in other studies more widely in the outer Firth of Forth area (Camphuysen et al., 2004; Kober et al. 2010, 2012; Lane et al. 2020). The majority of gannet within the Proposed Development area are likely to originate from Bass Rock and other nearby islands, whose colonies are collectively protected under Forth Islands Special Protection Area (SPA) (See Appendix 11.4). Gannet were not assessed in the Forth and Tay Marine Assessment 2020 but are thought to have increased by 33% in Scotland between 2003 and 2015 at an average rate of 2.9% per annum (Murray, Harris and Wanless, 2015).
- Guillemots are one of the most abundant seabird species in the northern hemisphere (JNCC, 2021c), and were the most abundant seabird species observed during baseline surveys at the Proposed Development (See Appendix 11.1). Overall, guillemot populations in Scotland are estimated to have increased in the last decade (JNCC, 2021c). According to the Forth and Tay Marine Assessment, guillemot populations have remained stable (Marine Scotland, 2020). The closest colonies to the Proposed Development, St Abb’s Head NNR (+5% to 42,905 individuals in 2018), Farne Islands (+103% to 64,042 individuals in 2019) and Fowlsheugh (+12% to 69,828 individuals in 2018), have all increased in population size since the 2000 counts (JNCC, 2021c). The majority of guillemot within the Proposed Development area are likely to originate from these colonies (See Appendix 12.4).
- The Firth of Forth supports one of the largest coastal roosting populations of herring gulls in Scotland and supports a significant population of lesser black-backed gull (NatureScot, 2020). The majority of individuals of both species from baseline surveys are likely to originate from the nearby Forth Islands SPA (See Appendix 12.4), as well as potentially from nearby towns and villages which gulls may use for nesting and foraging (Balmer et al., 2013; Rock and Vaughan, 2013). The number of birds nesting in these urban environments is not well known in the UK (JNCC, 2021d; JNCC, 2021e), although they may constitute approximately 8% and 17% of herring gull and lesser black back gull populations, respectively, based on Seabird2000 counts and additional data from national authorities (Calladine et al., 2006). According to the Forth and Tay Marine Assessment, the populations of both herring gull and lesser black-backed gull have remained stable during the assessment period (Marine Scotland, 2020) although herring gull populations in Scotland are estimated to have declined significantly since the Operation Seafarer census in 1969-70 (JNCC, 2021d; JNCC, 2021e).
- Kittiwake populations were assessed to have been stable between 2011 and 2018; however, their abundance is also estimated to be less than 50% of that recorded in the 1990s (Marine Scotland, 2020). Additionally, monitored colonies close to the Proposed Development have shown a decline since the Seabird 2000 census (Fowlsheugh -25% to 14,039 individuals in 2018; St Abbs Head NNR -58% to 4,651 individuals in 2019; Farne Islands -24% to 4,402 individuals in 2019; JNCC, 2021f). The majority of kittiwake within the Proposed Development area are likely to originate from these colonies (See Appendix 12.4).
- Puffin populations are overall estimated to have increased between the 1969-79 Operation Seafarer census and the Seabird 2000 census (1998-2002). However, between the Seabird 2000 census and the most recent counts in 2017/2018, the populations at the Isle of May and the Farne Islands are estimated to have declined by 43% to 39,200 apparently occupied burrows and by 21% to 43,753 apparently occupied burrows, respectively (JNCC, 2021g). The majority of puffin within the Proposed Development area are likely to have originated from these colonies (Appendix 12.4).
- The east coast of Scotland supports several large breeding colonies of razorbill. Razorbill populations are assessed to be increasing in Scotland (JNCC, 2021g), and within the Forth and Tay region ( Figure 3.2 Open ▸ ; Marine Scotland, 2020). One of the largest population increases since the Seabird 2000 census was at Fowlsheugh, where an increase of 121% was recorded in 2018 (JNCC, 2021g). The majority of razorbill within the Proposed Development area are likely to have originated from this colony (Appendix 12.4).
- In October 2021, a new strain of pathogenic avian influenza (HPAI) was identified in the UK. Originally recorded in domestic birds and poultry, it has more recently spread to seabird colonies around Europe. Infections have been recorded in several seabird species around the UK, including great skua Stercorarius skua, gannet, guillemot and kittiwake. The full extent of the outbreak upon seabird populations is currently unknown, as discussed in Section 5.4.
Figure 3.1 Breeding seabird relative abundance from 1986 to 2019. The dashed line shows the 95% confidence limits (NatureScot, 2018).
Figure 3.2 Summary of seabird status in the Forth and Tay from Scotland’s Marine Assessment 2011 - 2018 (Marine Scotland, 2020). Blue circle = some concerns, green square = few or no concerns, 3 stars shows that the underpinning data give high confidence.
4. Seabirds and the food web
- Trophic levels describe the hierarchal levels which organisms occupy in the food web. Primary producers, such as phytoplankton and seaweed, comprise the lowest trophic levels in marine food webs. They are consumed by primary consumers (herbivores) such as zooplankton, some crustaceans (e.g. copepods) and molluscs (e.g. clams, snails, mussels). Secondary consumers (carnivores or omnivores) such as fish larvae, herring Clupea harengus and lesser sandeel Ammodytes marinus (hereafter ‘sandeel’), and some crustaceans (e.g. crabs, shrimp) feed on primary consumers and primary producers. These species support tertiary consumers, including medium fish and cephalopods (e.g. octopus and squid species). Seabirds, along with marine mammals, large fish and elasmobranchs (sharks, skates and rays), are the top predators of the natural marine food chain. An example of a marine food chain which illustrates the interactions between the different trophic levels is presented in Figure 4.1 Open ▸ .
Figure 4.1 Significant interactions modelled between functional groups and drivers (from Lynam et al., 2017).
- Typically, the marine environment follows a ‘wasp-waist’ trophic structure, where mid-trophic level species have lower diversity, compared to high diversity in both high and low trophic levels. These mid-trophic level species play an important role in ecosystem functioning (Rice, 1995). In the North Sea, the main prey groups which dominate the mid-trophic level are planktivorous fish such as sandeel, herring, sprat, Norway pout and juvenile gadoid fish, all of which are founded within the Proposed Development area (see volume 2, chapter 9). These species link the lowest trophic levels to the highest (Mackinson and Daskalov, 2007; Fauchald et al., 2011; Lynam et al., 2017). Sandeel, sprat and herring are the most important prey species for many seabirds during chick-rearing in the North Sea, including puffin, razorbill, shag and kittiwake (Wanless et al, 2018).
- The North Sea is one of the most anthropogenically impacted marine ecosystems (Halpern et al., 2015; Emeis et al., 2015). Small, shoaling forage fish in mid-trophic levels experience top-down pressure from commercial fisheries, as illustrated in Figure 4.1 Open ▸ , whilst bottom-up processes driven by temperature have dominated changes to planktonic groups since the 1960s. These pressures propagate up and down the food chain, with mid-trophic fish linking the pressures on the upper and lower trophic levels ( Figure 4.1 Open ▸ ; Lynam et al., 2017). Monitoring seabird populations can therefore provide insight not only into the top-down pressures, but also into the bottom-up pressures such as climate change, and changes to fisheries, making them a good indicator of ecosystem health (Church et al., 2019; Ramos and Furness, 2022).
- Prey availability is one of the most important controls of species abundance and distribution in the higher trophic levels, including seabirds (Lynam et al., 2012; Mitchell et al., 2020). Reduced prey availability and changing prey distribution means that seabirds may have to forage further for food. For example, Fayet et al. (2021) compared the foraging costs in puffin populations in the north-east Atlantic. They found that puffins from declining populations in southern Iceland and north-west Norway had the greatest foraging ranges and least energy-dense diet. Low prey availability close to the colonies, potentially resulting from climate or commercial fisheries effects, is also amplified by increased intra-specific and inter-specific competition which forces birds to forage further from their colonies (Fayet et al. 2021).
- Diet and foraging behaviour determine the extent to which seabird species can respond to changing prey availability. Generalist species, such as gulls, which feed on a wide range of prey types will be more resilient to changing prey availability than more specialist species such as kittiwake which are surface feeders which predominantly prey on small fish (Furness and Tasker, 2000). Water column feeders, such as auks, forage from the surface to the seabed (depending on water depth) and can feed on both pelagic and demersal fish species, as well as invertebrates such as squid and zooplankton. Surface feeders, including kittiwake and terns, are restricted to prey available within the upper 1-2m of the sea surface, such as small fish, zooplankton and other invertebrates. Therefore, changes to prey distribution within the water column resulting from changes to stratification or temperature, for example, will affect surface feeding species differently to water column feeding species.
- This has been demonstrated in the North Sea, where almost 50% of surface feeding seabird species exhibited widespread breeding failures between 2010 and 2015; compared with only two of the eight-water column feeding species assessed ( Figure 4.2 Open ▸ ; OSPAR, 2017; Mitchell et al., 2018). Typically, seabirds that feed within the water column are better able to cope with changes in prey availability than surface feeding species, as explained above (Mitchell et al., 2020). This is likely to be linked to changes in the availability of small fish species (such as sandeel and sprat species) which are the predominant prey of surface feeding species such as kittiwake (Frederiksen et al., 2005; Carroll et al., 2017). A summary of the typical feeding strategy and prey of key seabird species for the Proposed Development has been outlined in Table 4.1. Plunge divers dive into the sea from a height to catch prey, whereas pursuit divers dive and can then swim underwater in pursuit of prey.
Figure 4.2 Proportion of seabird species (%) in the North Sea not showing widespread breeding failure, 1992 – 2015. The red horizontal line denotes the target threshold of 75 % or more. OSPAR, 2017)
- The availability of sandeels has been correlated with the breeding success and adult survival of kittiwakes (Frederiksen et al., 2004, 2008a; Carroll et al., 2017). Adult kittiwakes eat mostly older (1+ year group) sandeels during April and May; switching to juvenile (0-year group) sandeels in June and July during chick rearing (Lewis et al., 2001). This correlates with the annual cycle of sandeels. The 1+ year group (sandeels hatched prior to the current year) are active in the water column during spring. Once they have accumulated enough lipids they bury themselves in the sand, usually in June-July, and live off their stored lipids during the winter. The 0-year group (young of the year) sandeels are available from June onwards following metamorphosis from larvae into juveniles, and prior to burying themselves to overwinter (Wright and Bailey, 1996). However, density dependence also influences sandeel recruitment, and the biomass of the sandeel stock tends to be driven by occasional especially good years (ICES, 2017). In sandeel stocks with low fishing mortality, years with high stock biomass tend to show low recruitment, whereas high recruitment is more likely when adult stock biomass is lower (ICES, 2017, Lindegren et al. 2018). Both climate change and commercial fisheries are implicated with a reduction in sandeel abundance, which may contribute to kittiwake declines (Carroll et al., 2017).
- In the western North Sea between 1973 and 2015, the diet of chick-rearing kittiwakes, puffins, razorbills and shags was predominantly comprised of sandeels (Wanless, et al., 2018). Clupeids (sprat and herring) were the second-most important prey species, however these rarely exceeded 10% of the food biomass per year. Juvenile gadids were another important prey species (1 - 10% biomass) for these seabird species in some years (Wanless, et al., 2018).
- For guillemots, sandeels were the predominant prey until the late 1990s, when a shift to sprat (93%) and herring (7%) was observed (Wanless, et al., 2018). Between 1982 and 2019, sandeels were largely confined to the early part of the chick period as they have declined (Harris et al., 2022). A trend towards more sprat and herring have also been observed since the mid-2000s in razorbills and kittiwakes during chick-rearing, though sandeels are still the dominant prey (Wanless et al., 2018). Sprat feed and spawn repeatedly through spring and summer in coastal and offshore waters, and so are available for a wider period (MCCIP, 2018). Gannet predominantly feed on pelagic fish such as mackerel and sandeel, and also feed on fisheries discards, however the introduction of bans on discards may reduce the availability of this food source (Le Bot et al., 2019).
- Gull species, such as herring gull and lesser black-backed gull are able to feed on a diverse range of prey and food from both natural and anthropogenic sources (Table 4.1). In the south-eastern North Sea, faecal samples revealed that both lesser black-backed gulls and herring gull diets were predominantly composed of bivalves and crustaceans (Kubetzki and Garthe, 2003). A decline in herring gull abundance has been observed in Scotland since the 1969-70 National Census, and lesser black-backed gull populations have strongly fluctuated, which has been associated with changes in waste management such as covering refuse tips, and a reduction in fisheries discards (Burthe et al., 2014; Foster, Swann and Furness, 2017; JNCC, 2021d; JNCC, 2021e; Tyson et al., 2015); this may be evidence of the over-reliance of these species on these food sources. Foraging at landfills can also increase the risk of disease and mortality from Clostridium botulinum infection (Coulson, 2015).
5. Anthropogenic activities causing direct effects on seabirds
5.1. Climate change
- Climate change and severe weather events are one of the biggest threats to UK seabird populations (IPCC, 2014; Dias et al., 2019). Direct effects on seabirds which arise from climate change include storms, heavy rainfall events, flooding of nesting sites and temperature extremes.
- Strong winds and heavy rainfall during the breeding season can chill eggs, kill nestlings, and prevent adults foraging adequately to feed chicks, which can result in widespread breeding failures (Mallory et al., 2009; Mitchell et al., 2020). Flying and diving under high wind speeds can also impact body condition, by requiring greater energy expenditure (Kogure et al., 2016), and reducing foraging success. Storm conditions can reduce the availability of prey at the water surface and can increase water turbidity which reduces the ability of visual hunting seabirds, such as cormorants and shags, to find food. This can contribute to mass mortality events, which are further discussed in Section 6.4.
- Gannet may be able to take advantage of strong winds during adverse weather by modulating their flight height, giving them some resilience to storms (Lan, Spracklen and Hamer, 2019). For guillemot, the negative demographic impact of storms and adverse weather may be buffered to some extent by their large population size (Johnson et al., 2021).
- Seabirds are typically either ground nesting (e.g., herring gull, lesser black-backed gull, Arctic tern), burrowing (e.g., puffin), or cliff nesting (e.g. gannet, guillemot, kittiwake, razorbill). Flooding from heavy rain and storm surges can flood or wash away nests, and high winds can damage or destroy nests. Additionally, rising sea levels may reduce the habitat availability of low-lying nesting species, such as terns (Ratcliffe et al., 2008), with an estimated mean sea level increase of 4cm (likely range between 1 and 7cm) in 2018, when compared with the 1981-2000 average in the Forth and Tay region (Marine Scotland, 2020). For example, low productivity of Scotland’s puffins in 2007 has been partially attributed to unprecedented rainfall which flooded their burrows (Newell et al, 2013). An estimated reduction of 15% of kittiwake nests and 28.5% of razorbill nests was caused by a single storm in one year at the Isle of May (Newell et al., 2015). Gull species, such as herring gull and lesser black-backed gull, are able to utilise urban environments for nesting, which may give them resilience against any reduction in available nesting habitat (Balmer et al., 2013; Rock and Vaughan, 2013).
- Abnormally hot weather can cause stress to seabirds through issues with heat dissipation. For example, great skuas have been found to spend more time bathing in higher temperatures, meaning less time is spent foraging and tending to chicks (Oswald et al., 2008). Diving species may have significant heat loss to the water during diving, however this also generates metabolic heat (Oswald and Arnold, 2012). Puffins are thought to use their relatively large bills to dissipate excess metabolic heat, and so may be more resilient to hot weather than other species (Schraft, Whelan and Elliott, 2019). When combined with indirect effects of climate change, such as reduced prey availability, this can have critical effects on chick survival (Oswald et al., 2008; Hakkinen et al., 2022).
- There is a great degree of uncertainty in climate models for the North Sea, but there is potential that the magnitude and frequency of extreme weather events could increase as a result of climate change in the North Sea (Rahmstorf and Coumou, 2011, Wolf., Woolf and Bricheno, 2020). This would be likely to present population-level consequences for many seabird species, as outlined above (Frederiksen et al., 2008b; Louzao et al., 2019).
5.2. Commerical fisheries
- Commercial fisheries and seabirds have the potential to interact due to spatial and temporal overlap between foraging ranges and fishing activity (Žydelis et al., 2011; Lewison et al., 2014). In the vicinity of the Proposed Development, and in the wider Forth and Tay area, the dominant fishing types are demersal trawling, predominantly for Nephrops, followed by creeling for lobster and crab and dredging for scallops (volume 2 chapter 12; Marine Scotland, 2020).
- Bycatch (the incidental capture of seabirds by commercial fisheries) has significant impacts on seabirds globally (Tasker et al., 2000; Dias et al. 2019). Primarily, fishing gear in which seabirds are most likely to be bycaught are those involving fixed nets or longlines (Anderson et al., 2011; Žydelis et al., 2013). Annually, an estimated mortality of 400,000 birds is predicted to occur in gillnet fisheries (Žydelis et al., 2013) and an estimated hundreds of thousands in longline fisheries (Anderson et al., 2011). Whilst these fishing types are uncommon in the Forth and Tay, seabirds from the Proposed Development area may be exposed to these fisheries when foraging further offshore, or during migration.
- Bycatch from trawling can result from diving birds, such as guillemot, becoming entangled with trawls, or at the surface where seabirds can be struck by cables which are used to tow and monitor the equipment, thus injuring or drowning the bird (Løkkeborg, 2011). Commercial fisheries bycatch has been previously linked with a decline in population abundance in guillemot (Johnson et al., 2021).
- With commercial fishing effort increasing year-on-year (FAO, 2020), bycatch risk is likely to increase due to increased potential for interaction between industry and birds. Nearly 30% of seabird species analysed by Dias et al. (2019) were predicted to be affected by bycatch, with the number of globally threatened species affected increasing from 40 to 50 since 2010.
- Several British seabird species are at risk of being captured incidentally, with guillemots, razorbills and diver species being most susceptible (Bradbury et al., 2017; Larsen et al., 2021). Seabird foraging strategy affects susceptibility per gear type, with pursuit divers (e.g. guillemots) and plunge divers (e.g. gannets) more likely to be bycaught in gillnets rather than surface feeders such as kittiwakes (NatureScot, 2018). A study of Danish fishing boats operating within the North Sea found seabirds were more susceptible to bycatch in the winter compared to in the breeding season, potentially due to there being less prey available during winter, so seabirds come closer to the nets to secure food (Larsen et al., 2021).
- Recently, preliminary estimates of total annual seabird bycatch mortality in the UK have been calculated for 2016 and 2017, based on data from static net, longline and midwater trawl fisheries (Northridge, Kingston and Coram, 2020). Annually, 1,800-3,300 guillemots and 2,200-9,100 fulmars were estimated to be incidentally captured across the UK waters, predominately in static and longline fisheries, respectively ( Figure 5.1 Open ▸ ). However, true estimates may be higher than this as only UK vessels were included in the analysis (Northridge, Kingston and Coram, 2020). Demersal trawls, ring nets and pots were also not included in these analyses due to relatively low observation levels.
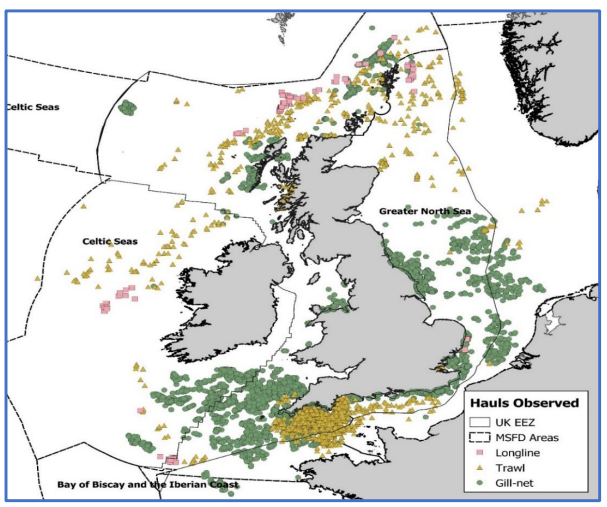
Figure 5.1 Observed hauls (left) and observed seabird bycatch by species (right) between 1996 and 2018 (Northridge, Kingston and Coram, 2020). Bycatch observed from 21,261 hauls across 3,455 trips in this period.
- Fisheries discards can also provide an alternative food source to generalist feeders, such as gulls and gannets, when natural prey species are not available, which may benefit some seabirds. However, since 2019 in the EU (European Commission, 2019) and the UK (Marine Management Organisation, 2019), landing obligations are being enforced which eliminates commercial fisheries discards, which will likely have reduced the availability of this food source (Uhlmann, Ulrich, and Kennelly, 2019; Le Bot et al., 2019), however the impact of this ban on seabirds is not yet known.
5.3. Offshore wind energy
- The potential for direct adverse effects on seabirds arising from the presence of offshore wind farms is broadly determined by overlap between species habitat and placement of wind farm arrays. The predominant direct effects acting on birds from the presence of offshore wind farms are related to collision, barrier effects and displacement (Gibson et al., 2017).
- Collision of seabirds with offshore wind farms may result in direct mortality or lethal injury (Desholm and Kahlert, 2005). The risk of collision varies between species and is affected by behaviour, population size, time of year, weather and wind farm design (Gill, 2005; Drewitt and Langston, 2006). Seabird species most at risk of collision with offshore wind farms are those which predominantly fly at the height of the wind turbine blades (Furness et al., 2013; Johnston et al., 2014). Eight seabird species present within the Proposed Development were considered to be vulnerable to collision based on the wind turbine height (Furness et al., 2013): kittiwake, herring gull, lesser black-backed gull, gannet, Arctic tern, common tern, little gull and great skua. Auks and fulmar were not considered vulnerable to collision as they generally fly at low altitudes, well below the minimum height of wind turbine rotor blades (Cook et al., 2012; Johnston et al., 2014; Jongbloed, 2016).
- For the Project, collision risk was assessed for kittiwake, herring gull, lesser black-backed gull, gannet, Arctic tern, and great skua (Appendix 11.3). For all species, the worst-case collision impacts were predicted for the smallest 14 MW wind turbine and the estimated number of annual collisions was highest for kittiwake. The consequences of collisions on seabird populations at SPAs are considered in Appendix 11.6. To reduce the collision risk to seabird species, Project design measures have been included, such as increasing the minimum lower blade tip height to 37m above LAT.
- Collision risk has been studied at a variety of offshore wind farms (Christensen et al., 2004; Blew et al., 2008; Krijgsveld et al., 2011; Leopold et al., 2013; Skov et al. 2018), however, studies reliant on carcass recovery at sea are inherently biased because carcasses drift, sink and can be scavenged. In offshore wind farms, models (e.g. Band, 2012) are used to predict estimated numbers of collisions for impact assessment purposes. Collision is more likely to occur if seabirds fail to avoid wind farms, although there is growing evidence of micro-, meso- and macro- scale avoidance, by a variety of species (Cook et al., 2014a; Bowgen and Cook, 2018).
- The presence of offshore wind farms can lead to displacement (where seabirds foraging within an area are displaced by the installation and operation of a wind farm) and barrier effects (where a movement corridor for seabirds has been effectively blocked by the installation of a wind farm) for some seabirds, with the potential for energetic consequences (Humphreys, Cook and Burton, 2015).
- Species which have been observed to exhibit displacement and barrier effects in northwest Europe include guillemot, razorbill, grebes, divers and gannets, whereas gull species may be attracted to the offshore wind farms (Dierschke et al., 2016). Displacement assessments were undertaken for kittiwake, guillemot, razorbill, puffin and gannet for the Project (Appendix 11.4) and the estimates of potential mortality due to displacement modelled for the relevant SPA populations in Appendix 11.6.
- Tracks of individual birds in the vicinity of three offshore wind farms at Helgoland, North Sea indicated that 89% of gannets avoided wind farm areas (Peshko et al., 2021). Analysis of GPS tracking data at the Proposed Development estimated that only 0.7% of the home range of gannet foraging from Bass Rock, the closest significant colony, and 74% of gannet did not enter the proposed development area at all (Appendix 11.4, Annex E).
- Increased energy expenditure associated with longer trip times between breeding colonies and foraging areas as a result of displacement and/or barrier effects can adversely affect local bird populations, with the potential to cause population-level effects (Masden et al., 2010). Displacement to other areas can also increase the pressure on mid-trophic prey species, such as sandeel, sprat and herring, in the areas which seabirds might be displaced to. For migratory birds, potential barrier effects can be reduced through altering wind turbine configuration to create corridors (Krijgsveld, 2014). However, the level at which species may be affected by barrier and displacement effects is difficult to quantify, with both likely to be confounded by other population-level effects such as fluctuations in prey distributions or the presence of fishing vessels immediately outside the wind farm boundary (Votier et al., 2010; Vanermen et al., 2013).
5.4. Avian influenza
- Avian influenza (commonly referred to as ‘avian flu’ or ‘bird flu’) is a virus that causes disease in birds, affecting the respiratory, digestive and/or nervous system of many species. Typically, infections are from a low pathogenic viral strain which causes mild illness. However, strains can mutate from low to high strains, which cause severe symptoms, often with high mortality rates and these strains may spread quickly causing an outbreak. The virus has become a disease of global significance due to poultry intensification creating conditions favourable for transmission of highly pathogenic strains (Gilbert and Xiao, 2008).
- In October 2021, a new strain of HPAI (H5N1) was identified in the UK. Since then, 120 further locations of infection in captive birds and poultry, have been identified across the UK, and 354 separate locations of infection across wild birds of 63 species have been identified across 76 countries worldwide (DEFRA, 2022). This has been the highest recent occurrence of HPAI in the UK with 90 cases of outbreak, compared with 28 in winter 2016/17 and 13 in 2020/21 (Lean et al., 2022). The greatest proportion of infection to date has been observed in swans, geese and ducks, and these species may form a natural reservoir of the virus. High occurrences of the 2021/22 avian flu have also been recorded in waders, gulls and auks as well as in species of birds of prey (DEFRA, 2022).
- In Scotland since October 2021, there have been HPAI (H5N1) infections recorded in several seabird species, including gannet at Bass Rock and Hermaness, Shetland (Martin, 2022) and Isle of Noss (Philip and Tyler, 2022), guillemot at St Abb’s Head (Hall, 2022) great skua at St Kilda, Fair Isle, Isle of Noss and Foula (Banyard et al., 2022; NatureScot, 2022a; Philip and Tyler, 2022) and kittiwake, great black-backed gull and terns at the Isle of May (Steel pers comm. 19 July 2022; NatureScot, 2022a). In August 2022, there had been no mass mortalities observed in Scottish tern, razorbill or puffin colonies (Philip and Tyler, 2022). However, the full magnitude of impact is currently highly uncertain.
- The effect of environmental variables on HPAI transmission and persistence is poorly understood (Gilbert et al., 2008; Lane et al., 2022). However, climate change is impacting the migration routes and distributions of birds, for example altering the timing of migration, stopover, and breeding, as well as the availability of breeding sites. This could lead to increased opportunity for transmission of HPAI between wild populations or between wild and domestic birds (Tian et al., 2014). Climatic changes may also alter the ability of the virus to survive in the environment, with potential to increase infection and transmission rates if there is longer environmental survival (Martin, Becker and Plowright, 2018).
5.5. Invasive non-native species
- Seabird populations have been severely impacted by the introduction of mammalian predators to regions where they would not ordinarily occur (Dias et al., 2019). Brown rats, cats, and American mink are among the introduced non-native predatory mammals to Scotland that pose a serious threat to seabird colonies. Native mammals brought by humans to offshore islands, such as hedgehog, stoat, and fox, also pose a significant threat (Marine Scotland, 2020). For example, American mink have been estimated to reduce herring gull breeding success by 41% in the west Scottish islands (Coulson, 2019). During site-specific surveys of the Proposed Development, no INNS were observed (see Appendix 22, Annex 22.8).
- Tree mallow is native to coastal areas of the Mediterranean and the south-west coast of the UK, but when introduced beyond these locations it often becomes invasive, causing significant changes to vegetation communities (Van der Wal, 2006). In the Firth of Forth region, declines in puffin populations have been linked to an increase in invasive tree mallow, which can block the entrances to puffin burrows and prevent breeding. Active management has led to a decline of the plant at Craigleith from 80% coverage to less than 10% between 2010 and 2020 (Anderson, 2021a; Anderson, 2021b).
5.6. Pollution
- Pollution in the marine environment which can affect seabirds includes oil pollution, other chemical contaminants, and marine litter such as plastics (Burthe et al., 2014). Oil spills can cause suffocation of seabirds and can disrupt feather integrity, leading to a loss of water proofing and buoyancy, and an inability to dive or fly (Troisi, Barton and Bexton, 2016). Chemical pollutants can accumulate in the tissues of predators such as seabirds, which can adversely affect survival rates and productivity (Letcher et al., 2010; Votier et al., 2005; Votier et al., 2011). For example, winter mortality of guillemot has been demonstrated to by doubled by major oil pollution incidents (Votier et al., 2005).
- Marine litter is persistent, manufactured and/or processed anthropogenic material found in the marine environment, the most common of which is plastics and fishing gear (UNEP, 2021). Between 2016 and 2020, plastic production in Europe increased by almost 10% (Plastics Europe, 2021), a proportion of which is likely to end up in the marine environment as plastic waste. After encountering marine litter, many marine species ultimately end up ingesting it or becoming entangled, which can be fatal (Good et al., 2010; Roman et al., 2019).
- The ingestion of marine litter can be detrimental by blocking or puncturing intestines and through the assimilation of harmful chemicals which leach out of items (Carey et al., 2011; Tanaka et al., 2013). Some models indicate 99% of all seabird species are predicted to ingest plastic by 2050 (Wilcox et al., 2015). One study indicated seabirds can have around a 20% chance of fatality from ingesting just one piece of marine litter, with the composition of the litter influencing probability of death (Roman et al., 2019). Some species groups are more susceptible to ingestion of marine litter due to their life history strategies, with surface feeders such as fulmars more susceptible than pursuit divers such as guillemots due to the assimilation of marine litter on the water surface (Provencher et al., 2010; Daunt and Mitchell, 2013). Smaller items such as microplastics may also release chemicals which are ingested by organisms in lower trophic levels and can bioaccumulate in high trophic level species, such as seabirds (Miller et al., 2020).
- When considered alongside climate change, the effects of marine litter can be made exponentially worse, with the potential for items to add to negative feedback loops through emission of greenhouse gas during the lifetime of items (Ford et al., 2022). Climate change also has the potential to affect marine litter by increasing flux into oceans through extreme weather events and associated run off, in addition to affecting degradation through heat and light exposure (Deng et al., 2021). Fragmentation of items and increased likelihood of small items remaining in suspension may also be more likely to occur under storm conditions, which are expected to increase due to climate change.